
An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Psychopharmacol Bull
- v.50(2); 2020 May 19

Case Report: A Case of Intellectual Disability with Inappropriate and Challenging Sexual Behavior that was Treated with GNRH Analogues
Intellectual Disability starts within the course of developmental stages and covers both intellectual and adaptive deficiencies in conceptual, social and applied fields. Individuals with intellectual disability experience many difficulties in social life due to challenging and inappropriate sexual behaviour. Suchdifficulties need to be addressed, reduced or treated. Traditional treatments often fail to treat and improve suchbehavior. Alternative treatment options need to be explored with studies conducted in this field. With this paper, we aimed to show and touch on alternative treatments for challenging and inappropriate behaviors of a 15-year old boy with intellectual disability, who was treated with GNRH analogues.
Introduction
Intellectual Disability is defined in DSM-5 as a conceptual, socially defective disorder that covers both intellectual and adaptive deficiencies and starts during the developmental stage. 1 Aggressive and challenging behaviors are frequently seen in these individuals with limited coping skills. 2 These behaviors, which are considered as the most important factors in applying to psychiatry clinics and drug use, make it difficult for caregivers that provide care to and live with intellectually disabled individuals. 3 , 4 Individuals with intellectual disabilities may have difficulty interpreting behavioral cues within their social environment. That is an obstacle for them to express their sexual desire in socially acceptable ways. 5 However; aggressive and challenging behaviors as well as inappropriate behaviors can be seen frequently in these individuals. These inappropriate behavior might be listed as masturbation, genital exposure, inappropriate sexual touching and embracing others too close. 6 – 8 Challenging and inappropriate sexual behaviors in individuals with intellectual disability are significant problems that need to be addressed in clinical practice. Antipsychotics are frequently used in clinical practice to manage these behaviors. However, there is a discrepancy regarding reports of the efficiency of antipsychotics medication in studies conducted in this field. 9 , 10 So, these drugs may not be efficient in the management of such behavior in certain individuals with intellectual disability. Despite ethical concerns, drugs that reduce sexual desire are sometimes used to reduce inappropriate sexual behavior. There are case reports that report GnRH analogues might be beneficial in cases with inappropriate sexual behavior and aggression in individuals with autism. 11 – 14 The use of GnRH analogues has several risks including delayed pubertal development, decreased muscle mass and bone density. In this case report, we aimed to discuss the use and efficacy of leuprolide as a GnRH analogue in a patient diagnosed with Intellectual Disability with challenging and inappropriate sexual behaviors that could not be suppressed by antipsychotics or other drugs.
A 15-year-old boy was brought to our child psychiatry clinic due to his aggressive and inappropriate sexual behaviors such as completely stripping off his clothes, and inappropriately touching private parts of the individuals in the household and others. It was noted that the patient had difficulties in speaking and language skills, interpersonal relations, academic and self-care skills as a result of the psychiatric examination. It was learned from the medical history that he was diagnosed with intellectual disability at primary school and he attended a special education center and a school where individuals with special needs went. He was asked to perform Wechsler Intelligence Scale for Children-Revised (WISC-R), bu he could not cooperate. According to the clinical opinion and history of the clinician and psychologist, he was diagnosed with moderate intellectual disability. There was no comorbid psychiatric diagnosis in psychiatric examination and history. It was learned from his parents that aggressive and inappropriate sexual behavior started in adolescence and has been on the increase for the past year. Also, It was learned that despite behavioral interventions and psychopharmacological treatments recommended by child psychiatrists they have met so far, no improvement was observed. Hormonal therapy was planned because the aggressive and inappropriate sexual behaviors persisted against all treatment interventions, it was noted that these behaviors made it difficult to live with the patient. The family was informed about the hormone treatment and associated effects and side effects. The patient was referred to a pediatric clinic for endocrinological and neurological examination. As a result of the physical examination; he was diagnosed with obesity (BMI: 36,26). Total testosterone level was 225 ng/dl. The diagnosis of Prader Willi Syndrome was excluded via genetic analysis. Consequently; it was concluded that GNRH analogue might be a treatment option. This alternative treatment option was used for reducing inappropriate and challenging sexual behavior despite ethical concerns, according to previous case reports. 11 – 14
Parents wereinformed about the uncertainties including costs, treatment expectations and possible side effects related to off-label use and written consent was obtained from the parents. 3.75 mg leuprolide acetate was administered intramuscularly once in every 28 days. No other psychotropic drug was used simultaneously. The patient was examined once a month for possible side effects and puberty evaluation. Strength and Difficulties Questionnaire (SDQ) and Turkish version of Aberrant Behavior Checklist (ABC) were completed by parents at the onset of treatment and every following month. It was learned that the boy’s challenging and inappropriate sexual behaviors increased after the first and second doses of leuprolide. There were no decrease in SDQ ve ABC scores. Parents’ anxiety gradually increased due to worsening of the symptoms. After the third dose, he was admitted to the psychiatry inpatient unit for observation. No other medication was prescribed, other than lorazepam that was to be used whenever required. During his 2-week inpatient stay, his challenging and inappropriate sexual behaviors increased gradually. Therefore leuprolide treatment was stopped. Testoterone level was 270 ng/dl after the third dose. SDQ and ABC scores were presented in Table1 .
Challenging and inappropriate sexual behaviors in adolescents with intellectual disability may cause significant concerns for others and restrictions for the individual. There may also be more serious negative consequences for these individuals, such as forensic events. Despite ethical concerns, hormonal treatment options are sometimes considered especially in the management of inappropriate sexual behaviors.
Although there is limited literature on the use of GnRH analogue in management of challenging and inappropriate sexual behaviors, results from individuals diagnosed with autism spectrum disorder promise that this treatment option might be beneficial. 11 – 14 In some case reports; GnRH analogue therapy has been reported to reduce inappropriate sexual behavior. 11 , 13 , 14 Robin et al. (2016) reported that GnRH analogues were effective in managing such behavior in a male patient with autism, who had severe aggressive behavior since the onset of puberty. 12 A positive effect of combined long acting GNRH agonist treatment and heavy metal chelation therapy on aggressive behavior has been reported in 11 autistic children. 15 , 16 Based on these previously reported clinical experiences, we decided to use a GnRH analogue to manage and reduce challenging and inappropriate sexual behavior in this patient with Intellectual Disability. Another reason for this decision was that these problems did not decrease with behavioral methods, education and commonly used pharmacological drugs. 3.75 mg leuprolide acetate was administered intramuscularly and 3 doses every 28 days. Our aim was to reduce gonodal steroid levels to pre-pubertal levels. However, during the follow-up, we observed that the patient could not tolerate the treatment and there was an increase in both challenging and inappropriate sexual behaviors. There were no changes in SDQ scores and increases in ABC scores. Therefore, treatment with leuprolide acetate was stopped.
It is known that administration of leuprolide acetate in humans initially causes a temporary increase in gonadal steroid levels (testosterone and dihydrotestosterone in men). A mild increase in testosterone levels was observed in our patient after the administration of leuprolide acetate. Although there was a mild increase in testosterone levels, we thought there might be a difference in testosterone receptor sensitivity. The negative outcome of treatment in this case is different from the positive experiences mentioned in the literature. It should be noted that GnRH analogue therapy might not be beneficial and efficient in all patients. We believe that this case report will make a significant contribution to the limited literature on the use of GnRH analogues in challenging and inappropriate sexual behavior.
Case Study of a Child with Intellectual Disability
In this intellectual disability case study, the author looks at designing an education curriculum for Meagan, a 14-year-old student.
Introduction
There are numerous interventions that have been designed to enable students with intellectual disability better cope with their condition. Most of these interventions have been hinged on the principle that respective educational programs should be tailored to complement the students’ strengths, and to supplement their weaknesses.
This is the same principle underlying the functioning of the K12 educational principle because it is centered on meeting individual student needs (K12 Inc. 2011, p. 1). The K12 educational paradigm mostly works through online communication but it has been seen to offer a lot of advantages to students with unique disabilities.
For instance, the educational methodology is known to provide rich, challenging and engaging content; an individualized learning plan; a learning coach; and cutting-edge technology in the provision of the best learning outcomes for intellectually disabled students (K12 Inc. 2011, p. 1).
When educating students with intellectual disabilities, it should be understood that, students are bound to have trouble in learning, retaining information and understanding information (Pearson Education Inc. 2011, p. 2).
Educators have often experienced such challenges, but comprehensively, there is a consensus among most stakeholders that it is vital to make accommodations for certain groups of students, and it is also crucial to make curriculum modifications for other students. In this regard, there seems to be a lack of consensus in coming up with one formula for handling students with intellectual disability.
This is the main framework for the advancement of this paper because this paper focuses on developing curriculum adjustments for a young man, Meagan. Meagan is 14 years old and has had a history of intellectual disability. This paper analyzes various dynamics of Meagan’s life, with the intention of making curriculum adjustments to provide an effective framework for learning.
To provide a good backdrop for the development of a good framework for learning, several aspects of Meagan’s life will be analyzed. These aspects include his family background, personal history, personal skills and personal abilities.
These factors will be analyzed systematically. Comprehensively, this analysis will be done with the aim of identifying one long-term aim or objective of the learning outcome and two short-term aims or objectives of the learning outcome.
Family Background
Meagan is the eldest child in a family of three children. His younger sibling is a girl, Sophia, aged nine years old. The youngest child is also a girl and she is three years old. Among his siblings, Meagan is deemed to be the child who has experienced most difficulty in learning. Meagan’s family hails from a middle-class society in Melbourne, Australia. His father works as a retired engineer in a local factory.
The mother works as a librarian in a local university. There has been no vivid or confirmed reports of intellectual disability among any of Meagan’s family members, though there have been unconfirmed reports of mental illness among some of Meagan’s relatives hailing from his father’s side of the family.
His aunt is said to experience occasional episodes of mental instability. However, there have been no confirmed reports of mental illnesses or cognitive disability from any of the family members of Megan’s mother.
Megan’s family professes the Christian faith, though they are not committed in their religion. However, Christianity has had an influence on Meagan’s life because he strongly identifies with his Christian faith. In the past couple of months, Meagan was baptized and currently devotes most of his time to his religious duties. None of Meagan’s family members pay much attention to religion.
His family also hails from a background of child neglect, with many of Meagan’s relatives having been abandoned by their parents at an early age. Meagan’s parents are no exception. The degree of attention they give Meagan is inadequate because little attention is paid to Meagan’s slow intellectual development. This has been going on since his parents confirmed that he was suffering from intellectual disability.
There is also an almost non-existent family support structure for Meagan to cope with his condition. Moreover, there is very little evidence of family cohesion among Megan’s family members, starting from his parents to his siblings. In this regard, Meagan is left to live with his condition, alone.
Personal History
Meagan hails from the aboriginal community of Australia. He was prematurely born because he was birthed at only seven months into his mother’s pregnancy. During his infant life, Meagan was abandoned by his mother, even before he was completely weaned from her. This forced his father to look for a baby sitter.
Nonetheless, despite these challenges, Meagan lived to have a vibrant childhood, with no signs of failing to cope with his playmates or friends. To a large extent, Meagan has been deemed a “normal” child. In his teen years, he used to participate in church activities (for the young) and also took part in school activities including extracurricular games.
He was a vibrant member of the school choir and an active member of the school soccer club. However, Meagan’s repeated the seventh grade level (twice) because he failed to meet the minimum threshold for admission into the eighth grade.
For a long time, he experienced a lot of difficulty trying to meet the minimum threshold for admission into sequential class grades because he always trailed among the last five candidates in any class. This was witnessed from his admission into the first grade.
However, Meagan’s academic background was characterized by exemplary performance in various academic writing competitions. His teachers termed him as a very creative writer and he never disappointed in his English creative writing assignments.
However, this was as far as his academic excellence stretched. Currently, Meagan undertakes blue collar jobs on minimum wage but there is increasing pressure among his peers for him to continue with his studies.
Personal Skills and Abilities
Meagan has a creative mind. He has shown interest in creative writing from his younger years but as he grew older, his interest changed. However, as explained in earlier sections of this study, in his young years, Meagan used to write exemplary creative works. His interest however shifted into music when he grew a little older.
So far, he has been able to record music in a local music company but his talents have never been fully exploited because of the lack of adequate finances to bankroll his musical ambitions. Moreover, there has been limited support from most of his family members in his quest to pursue music. However, due to his strong religious background, Meagan hopes to produce music for his local church.
The main aim of undertaking a curriculum adjustment for Meagan is to enable him to earnest his abilities and use them to the optimum benefit of his talents.
To enable Meagan to be independent and able to communicate his needs in effective and acceptable ways.
To assist Meagan to excel in personal growth and compete with other students in varying levels of excellence.
Curriculum Adjustments
Making the best curriculum adjustments for Meagan entirely depends on the nature of his disability. From previous sections of this paper, we have affirmed that Meagan suffers from a slow comprehension of academic disciplines, but he has a stronger grasp on creative works.
Here, there are several curriculum adjustments that can be done to ensure Meagan lives to his full potential. In this regard, this paper proposes several curriculum adjustments, based on the K12 teaching model which aims to provide individualized learning for students with intellectual disability. They are outlined below:
Interest and Student Ability
To ensure Meagan lives to his full potential, it is crucial to make curriculum adjustments to suit individual needs, abilities and preferences. A uniform curriculum which is meant to work for the majority student population is bound to fail for Meagan because it will not be specific to Meagan’s abilities and potential.
In this regard, it is therefore crucial for the curriculum to be designed to emphasize on creative works, as opposed to academic excellence, to enable Meagan to succeed in arts (Queensland Government 2011). Emphasis should be further made to ensure the school grading criteria focuses the same level of attention it gives to sciences (and other disciplines) as it does with art subjects.
Such a grading criterion would ensure students are assessed on all fronts, and not just academic. When adjusting the learning curriculum, it is also crucial for teachers to structure the curriculum in a manner that guarantees the grouping of students into different ability groups.
Not all students have the same type of abilities and therefore, it would be beneficial for teachers to group Meagan into the “creative works” group, so that he can share his creative ideas with his peers (Foreman 2009, p. 170).
Adjusting the Learning Outcomes
Adjusting the learning outcomes is an important adjustment to the learning curriculum if the school grading process is to be fair. Here, “fair” means to accommodate intellectually disabled students (Snowman 2011).
Accommodation of Diverse learning Styles
Intellectually disabled students are normally faced with the challenge of failing to comprehend learning instructions as fast as other students do. However, research studies affirm that some of these students prefer certain learning styles in place of others (Queensland Government 2011). Moreover, educationists have shown that certain learning styles are more effective for intellectually disabled students, while others are not.
Such dynamics withstanding, it is crucial to make curriculum adjustments that allow for the accommodation of diverse learning styles for improved efficacy in learning. For instance, conventional or online lessons can be administered using various learning materials such as DVDs, CDs, Books, videos and such materials (Browder 2011, p. 332).
The inclusion of such diverse strategies is set to improve the level of interaction between the students and the teachers because an appropriate learning style would motivate the students to pay more interest in the learning process. This improves the students’ level of engagement. Moreover, such curriculum changes ensure the learning process is rich in its contents.
Integrating a Learning Coach (Parent Involvement)
It is crucial to integrate the input of a learning coach into the school curriculum to encourage the participation of Meagan’s parents in his educational endeavors. The parents will be the learning support team.
Already, we have established that Meagan hails from a family that pays little attention to his educational needs. Here, there is a strong need to integrate the parents’ input into Meagan’s educational projects to ensure he enjoys a support structure, aside from the traditional teacher-student framework.
Though an integration of the role of the learning coach into the school curriculum may not necessarily be confined in the parent-student framework, it is crucial for this integration to be developed in this framework, if Meagan has to develop better learning skills (National Parent Teacher Association 2009, p. 1 ) .
This is because a great degree of the deterioration of his intellectual ability comes from a lack of effective support structure that enables him to improve his learning skills (Queensland Government 2011).
For long, this need has been ignored, and as a result, Meagan has continually performed poorly in his academic endeavors. Nonetheless, the learning coach framework can be designed in various ways. For instance, the school curriculum can be designed to include the participation of parents in the student’s projects, at least once or twice a semester.
Parents may be required to give consent, provide counsel or similar activities on the student’s tasks, thereby encouraging him to better develop with his learning activities. The inclusion of this principle into the school curriculum may be indirectly beneficial to Meagan because it is bound to have a motivating effect on him. This is the first strategy that can be adopted in encouraging parent participation.
The second strategy that can be adopted by the school is implementing a family-school partnership policy where parents and teachers agree on a common framework where parental involvement is assessed, and the parents’ progress is measured (Westwood 2011, p. 15).
This recommendation emanates from research studies which have shown that schools which have an efficient family-school partnership perform better than schools which lack this policy (Queensland Government 2011).
Finally, the school should make adjustments to the curriculum to ensure that parents take part in the decision making process of activities affecting student achievement. Here, parents should be allowed to be part of advisory committees which affect student achievement.
This paper proposes that, adjustments in the school curriculum which have to be made to accommodate Meagan’s skills and abilities have to be done within the confines of earnesting his skills and abilities (to use them for the benefit of his personal growth). In this regard, this paper proposes that the school curriculum should be tailored to accommodate Meagan’s artistic skills.
Moreover, the learning outcome should be adjusted to accommodate the same skills and abilities. From a holistic perspective, this paper also proposes that diverse learning styles should be accommodated into the learning curriculum to ensure students with intellectual disability learn in an efficient way.
These recommendations are carved from the K12program. Nonetheless, this paper also puts a lot of emphasis on the importance of incorporating parent input in the school curriculum. Integrating these principles will go a long way in enabling Meagan to earnest his strengths and use them to the optimum benefit of his talents.
Browder, D. (2011) Teaching Students with Moderate and Severe Disabilities . New York, Guilford Press.
Foreman, P. (2009) Education of Students with an Intellectual Disability: Research and Practice (PB). New York, IAP.
K12 Inc. (2011) How a K12 Education Works . Web.
National Parent Teacher Association. (2009 ) Enhancing Parent Involvement. Web.
Pearson Education Inc. (2011) Teaching Students with Special Needs . Web.
Queensland Government. (2011) Intellectual Impairment – Educational Adjustments. Web.
Snowman, J. (2011) Psychology Applied to Teaching . London, Cengage Learning.
Westwood, P. (2011) Commonsense Methods for Children with Special Educational Needs . London, Taylor & Francis.
- Chicago (A-D)
- Chicago (N-B)
IvyPanda. (2023, October 29). Case Study of a Child with Intellectual Disability. https://ivypanda.com/essays/case-study-of-a-student-with-intellectual-disabilities-essay/
"Case Study of a Child with Intellectual Disability." IvyPanda , 29 Oct. 2023, ivypanda.com/essays/case-study-of-a-student-with-intellectual-disabilities-essay/.
IvyPanda . (2023) 'Case Study of a Child with Intellectual Disability'. 29 October.
IvyPanda . 2023. "Case Study of a Child with Intellectual Disability." October 29, 2023. https://ivypanda.com/essays/case-study-of-a-student-with-intellectual-disabilities-essay/.
1. IvyPanda . "Case Study of a Child with Intellectual Disability." October 29, 2023. https://ivypanda.com/essays/case-study-of-a-student-with-intellectual-disabilities-essay/.
Bibliography
IvyPanda . "Case Study of a Child with Intellectual Disability." October 29, 2023. https://ivypanda.com/essays/case-study-of-a-student-with-intellectual-disabilities-essay/.
- Relationship Between Language and Thought
- Coaches’ Performance in Educational Institutions
- General Curriculum for Intellectual and Developmental Disabilities Learners
- Influence of Coaches on Players
- Students With Intellectual Disabilities and Their Independence
- Intellectual Disabilities and Limitations for Human Life
- Leadership: A Tale of Two Coaches
- Intellectual Disabilities and Higher Education
- High Incidence Disabilities and Pedagogical Strategies for Learning Disabilities
- How Safe Are Your Kids With Volunteer Coaches?
- IEP Analysis
- Students with Asperger syndrome
- Classroom Design for Children With Disabilities
- Impact of Individuals With Disability Education Act (IDEA) on Education
- Annual Educational Goal for Students with Autistic Spectrum Disorders
Academia.edu no longer supports Internet Explorer.
To browse Academia.edu and the wider internet faster and more securely, please take a few seconds to upgrade your browser .
Enter the email address you signed up with and we'll email you a reset link.
- We're Hiring!
- Help Center

Case Study: Accommodating Students with Intellectual Disabilities

Students with intellectual needs have problems with their cognitive capability and usually lack skills in social behavior and self-care (Jones & Goble, 2012). Even though there is a wide assortment in the cognitive ability of individuals with intellectual disabilities, they generally have an IQ of 70-75 or even lower (Gordon, 2018). For students who are diagnosed with a disability, learning is usually faced with numerous challenges. Some of these challenges include being excluded from classrooms, being left out of field trips and lacking enough staff to help them learn (Hartmann, 2015). Despite their entitlement to inclusive education, students who have intellectual disabilities face academic and social barriers, which exclude them and makes them vulnerable to bullying (Gordon, 2018). The outcomes paint a stark picture of the way the education system fails to accommodate these individuals. There is a need to accommodate these students in schools so that they receive the equitable quality of education they deserve (Jones & Goble, 2012). Schools need to make adjustments on the way information can be presented to these individuals so that they can better engage in learning (Massouti, 2018). This report offers an analysis of the nature of inclusion, inclusive schools and suggests strategies to support and promote accommodation to students with intellectual needs.
Related Papers
Gillian Parekh
The framing of disability is an ongoing, negotiated discourse in which participants build upon, challenge, and reject the political, social, economic, and cultural influences that lead to constructions of impairment. Experiences of racialization, poverty, immigration, gender, and sexuality juxtaposed against defined institutionalized norms and dominant narratives speak to how disability is not only conceived but also experienced. Drawing upon transnational and citizenship theory, this thesis proposes employing a new framework of analysis, centralizing the experience of social citizenship and belonging as an indicator of broader structural equity. Situated in the field of education, theoretical considerations also explore how growing market fundamentalism shapes public schools and contributes to the systematic exclusion of poor and racialized students through mechanisms of disablement such as reduced academic programs and special education placement. This body of work includes three separate, but related, studies exploring historical and current incidences of institutional exclusion. In particular, the nuanced relationship of exclusion to race, class, gender, generational status, and sexuality, complicated with the identification of impairment, is explored. One of the most profound findings of this research is that, although there is much discussion in Disability Studies of the construction of impairment labels, this is the first quantitative analysis to substantiate these claims. Results also indicate that the classroom represents the most stratified space in which student groups defined by race, exceptionality, class, and generational status experience the greatest sense of exclusion. Evidence shows that employing a lens of citizenship and belonging is an authoritative tool in identifying the existence of inequities distributed among myriad identity groups. Furthermore, evidence lends credence to the notion that identification of disability is intimately linked to race, gender, and class contexts.
SamJoey Sam
Robert S. Brown
Cameron Crawford
Chris K . Chang-Bacon
This research report documents the nature and impact of a Consortium of 10 school boards a liated with the Council of Ontario Directors of Education (CODE) in leading bold and sophisticated change for today’s students, in one of the highest performing and most culturally diverse educational systems in the world – the province of Ontario in Canada. Over more than a decade, these boards built on an earlier approach by all of the province’s 72 boards to advance deep learning not by imposing reform from the top, or by supporting a multitude of initiatives among teachers and schools at the bottom of the system, by what educators themselves describe as “Leading from the Middle” (LfM).
Exceptionality Education …
Debra Pepler
Inquiry into practice
Saskia Van Viegen
International Journal of Disability, Development and Education
Meghann Proulx
Michelle Villeneuve
Despite recognition of the importance of parent involvement to enable meaningful inclusion of young children with developmental disabilities in education contexts, few Canadian studies have reported how parents experience this collaboration. Recent research suggests that the transition process is critical to the early school experiences of children with developmental disabilities yet challenging for their parents. The purpose of this paper is to report preliminary data from HELPS Inc, a Canadian multi-method research project describing Health, Education, and Learning Partnerships Promoting Social Inclusion of young children with developmental disabilities. In this paper we present parents’ perspectives on inclusion and collaboration with case study findings based on the experiences of three families in one Ontario community. The data demonstrate the unique experiences of and meanings of collaboration held by individual families, and highlight the challenges these differences pose fo...
RELATED PAPERS
Patricia Rice Doran
Exceptionality Education International
Judy Lupart
Cynthia Webb
Isabel Killoran , Gillian Parekh , Dustin N Graham
Isabel Killoran
International Journal of Inclusive Education
Kathryn Underwood
Kelly Gallagher-Mackay
Terri-Jane Stapleton
Vanessa Hamilton, OCT
W. John Williamson
Forum for International Research in Education (FIRE) Special Issue, Religious Identity and Education: A Response to Contemporary Global Pressures Vol 5, No 1
W. Y. Alice Chan , Harriet Akanmori , Crystena A . H . Parker-Shandal
Kim Calder Stegemann , Nan Stevens
Nicholas Ng-A-Fook
Kimberly Maich
Shirley Kendrick , Nadya Tymochenko
Journal of Bisexuality
Margaret Robinson
Mimi Masson
Benjamin Kutsyuruba , Allison Segeren
College of Alberta School Superintendents (CASS)
Barret Weber
Kimberly Maich , Carmen Hall
Edmore Mutekwe , Andrea Versuti
Mayumi Terano
Marcea L Ingersoll
Lynette Shultz
Kimberly Maich , Steve Sider
Maren Elfert , Lynette Shultz
State of French Second Language Education in Canada 2017
In M. Chitiyo, G. Prater, L. Aylward, G. Chitiyo, E. Dalton, & A. Hughs (Eds.)
Carla Reis Abreu-Ellis
Michelle Servais
Margo Greenwood
Panayota Gounari , Journal of Pedagogy , Γιώργος Γρόλλιος
Peter Grootenboer , Levon Blue
This paper has not been submitted for publishing.
Debora Oliveira de Lima
Intervention in School and Clinic
Kimberly Maich , Christina Belcher
Susan Drake
Education Watch
Gabrielle Young , Jeffrey MacCormack
RELATED TOPICS
- We're Hiring!
- Help Center
- Find new research papers in:
- Health Sciences
- Earth Sciences
- Cognitive Science
- Mathematics
- Computer Science
- Academia ©2024
- Research article
- Open access
- Published: 28 May 2016
Crime and victimisation in people with intellectual disability: a case linkage study
- Billy C. Fogden 1 ,
- Stuart D. M. Thomas 1 , 2 , 4 ,
- Michael Daffern 3 &
- James R. P. Ogloff 3 , 4
BMC Psychiatry volume 16 , Article number: 170 ( 2016 ) Cite this article
80k Accesses
57 Citations
38 Altmetric
Metrics details
Studies have suggested that people with intellectual disability are disproportionately involved in crime both as perpetrators and victims.
A case linkage design used three Australian contact-level databases, from disability services, public mental health services and police records. Rates of contact, and official records of victimisation and criminal charges were compared to those in a community sample without intellectual disability.
Although people with intellectual disability were significantly less likely to have an official record of victimisation and offending overall, their rates of violent and sexual victimisation and offending were significantly higher. The presence of comorbid mental illness considerably increased the likelihood of victimisation and offending; several sex differences were also noted.
Conclusions
People with intellectual disability are at increased risk for both violent and sexual victimisation and offending. The presence of comorbid mental illness aggravates the risk of offending and victimisation. Future research should focus on a more nuanced exploration of the risks associated with intellectual disability and specific mental disorders and related indices of complexity.
Peer Review reports
People with an intellectual disability (ID) are a marginalised and vulnerable group. The available research suggests an association between ID and criminal offending [ 1 – 3 ]; this has served to propel public fear and reinforce perceptions of the need for social distance. However, the evidence from which these conclusions have been drawn remains far from definitive, with significant methodological limitations marring what are arguably tentative conclusions [ 1 , 2 , 4 ]. A related area that has received much less scientific attention is criminal victimisation, despite a compelling argument that specific deficits in interpersonal functioning and cognitive capability potentially increase exposure to dangerous situations, therefore contributing to the likelihood of criminal victimisation [ 5 – 14 ].
Intellectual disability and criminal victimisation
Intellectual Disability is characterised by significant impairments in intellectual functioning alongside difficulties in daily tasks, personal responsibility and communication [ 15 , 16 ]. From a theoretical standpoint, Routine Activities Theory [ 17 ] conceptualises victimisation in relation to an interaction between an available victim, the absence of a capable guardian, and a motivated offender. It reasons that people with similar lifestyles or routine activities face similar victimisation risks as they are exposed to risky places and potential offenders [ 5 ]. Some research has suggested that people with ID are most commonly victimised by their carers [ 5 ]. Routine Activities Theory would argue that victims are easily accessible in their home / living environment (availability) and there is less protection of the victim if the perpetrator is the guardian (absence of a capable guardian); the carer offender may be motivated to offend due to carer stress, a provocative or frustrating incident, in this context offending may be facilitated by increased potential to evade prosecution (motivated offender). A study by Sobsey [ 18 ] supports this proposition and demonstrates the substantial vulnerability of victims with ID, noting that 44 % of perpetrators had contact with a victim through disability services, in which the victim was in close proximity with the perpetrator and was less likely to recognise or report a crime due to the apparent legitimacy of the disability service. As such, victimisation can be seen as a product of complex interactions between the environment, the victim(s) and the perpetrator(s).
The available research is consistent with this theoretical stance, noting increased rates of victimisation among people with ID compared to the general population. Wilson and Brewer [ 6 ] estimated that a diagnosis of ID doubled the risk for victimisation and vastly increased the likelihood of sexual assault and being the victim of robbery compared to the general community. Other studies report similar findings, estimating that the risk of victimisation is between three and seven times higher in people with ID compared to the general community [ 11 , 13 , 19 ]. Of particular concern are the high rates of sexual victimisation [ 19 – 22 ] and the finding that the majority of crimes were committed multiple times [ 23 ], across a range of ages and environments [ 13 , 24 ]. Of note, these victimisation experiences are rarely reported to authorities [ 6 , 7 , 19 , 23 ].
Surprisingly, there is little knowledge on the relationship between gender and victimisation among people with ID. While there is some evidence from general population estimates implicating males as being more vulnerable to victimisation [ 25 ], it is not known whether this trend exists in the ID population. That being said, tentative conclusions can be drawn from general disability research, which suggests that females are more susceptible to violent and sexual victimisation compared to males [ 20 , 26 ]. In sum, the available literature suggests that people with ID are at a greater risk of victimisation compared to non-disabled members of the general public, however it is far from a robust conclusion. This is for two reasons; first, of the few studies conducted, too many focus on outdated retrospective self-report data, which rely on the individual recalling events and judging whether a crime was committed. This presents a difficulty with people who have an ID as there are often disruptions in their memory functioning and judgements about their own, and the activities of others, when recalling crimes [ 7 ], consequently past research may either under report or over report actual rates of victimisation. Second, the operational definition of ID varies between studies and importantly, samples tend to be small and lack community comparison groups; both of which would arguably help contextualise the nature, direction and extent of victimisation [ 10 ].
Intellectual disability and offending
Like victimisation, the functional deficits evident in ID suggest that people with ID may also be likely to offend. This sentiment has a long tradition [ 27 ], attracting consistent research attention over the years; with studies claiming that people with ID are overrepresented among individuals processed by the criminal justice system [ 28 – 30 ]. The estimated prevalence of offending in people with intellectual disability ranges from two to ten per cent and varies depending on the population and methods utilised [ 27 , 31 ]. There is much variation within prison populations, with estimates ranging from less than 2 % to as high as 30 % [ 29 , 30 ], yet there is little agreement on a standardised conceptual definition of what criminal offending is across these studies. A recent systematic review, pooling results from ten studies and including a total of 11,969 prisoners concluded that typically 0.5 % to 1.5 % of prisoners are diagnosed with intellectual disabilities [ 32 ]. Estimating offending prevalence with prison populations is problematic as many individuals with ID have been diverted into the community or forensic services rather than prison, so there may be an under-estimation of the true prevalence using this method.
Court appearances and police contacts provide an alternative means of establishing prevalence and are more sensitive, as these records more adequately capture the extent of contact people have with the criminal justice system. The available literature at this interface estimates that around 1 in 10 people with ID will come into contact with the police or courts as a perpetrator of crime [ 14 , 33 ]. These rates are substantially different to those in the general population, with males with ID being three times more likely than males in the general public to have a prior conviction, while females have been found to be four times more likely to have a prior conviction. Interestingly, this figure was more pronounced for violent offences, with males four times higher and females 25 times higher, therefore potentially suggesting a significant vulnerability to violent offending among people with ID [ 34 ].
Some evidence suggests that people with ID are susceptible to the perpetration of specific crimes, such as sexual offences [ 2 ]. Further, there are additional factors that potentially complicate the hypothesised link between ID and offending, with findings revealing that complexities such as childhood neglect, physical health problems, mental health problems and perinatal adversity are particularly common among offenders with ID. There is also some suggestion that offenders with ID may be less effective at evading police and more visible as perpetrators [ 5 ] and this is the reason for increased prevalence rates.
The relationship between crime perpetration and ID and mental illness, which is highly comorbid with ID [ 33 ], has received empirical scrutiny. Hodgins and others [ 34 ] estimated that the presence of mental illness increased offending by five times in psychiatric inpatients compared to those with ID who had not been admitted for mental health treatment. Additionally, Vanny [ 33 ] found that nearly half of those people with ID who were referred to court had a mental illness, thereby suggesting a more complex group who may be at increased risk of criminal offending.
Aims and hypotheses
Against this background, this study sought to determine the prevalence of criminal victimisation and offending in an ID population and to compare this to a sample of people drawn from the general population. Based on the findings of Wilson and Brewer [ 6 ] it was hypothesised that people with ID would have higher rates of victimisation and offending relative to the community sample. Secondly, based on the findings by Sobsey [ 19 ], sexual crimes were expected to be increasingly more common in the ID group. Thirdly, the added complexity brought about by having comorbid mental illness [ 34 ] was hypothesised to increase the risks of victimisation and offending above that observed in people with ID only. Finally, males with ID were hypothesized to be at greater risk of both victimisation and offending than females with ID.
The study employed a case linkage design to compare rates of criminal victimisation and offending (operationalized as criminal charges) between those with a diagnosis of intellectual disability and a community comparison sample. The research complied with contemporary Australian National Health and Medical Research Committee (NHMRC) guidelines for conducting epidemiological research; administrative permission was granted for access to data stored on contact-based databases (see below).
Participant-level data were gathered through three archived databases; the Restrictive Intervention Data System (RIDS), the Victorian Psychiatric Case Register (VPCR) and the Victoria Police Law Enforcement Assistance Program (LEAP). The RIDS database is a state wide reporting system for individuals with a disability who have received a routine, pro re nata, or emergency restrictive intervention as defined in the Disability Act 2006 (Vic) . Section 3 of the Act defines ID as the concurrent existence of a significant sub-average general intellectual functioning, and significant deficits in adaptive behaviour, both which become manifest before the age of 18 years. Under the Act , a restrictive intervention can include chemical restraint, mechanical restraint or seclusion; these are mandatorily reported to Disability Services and recorded on the RIDS database and can only be used when they represent the least restrictive option. Episodes are updated monthly; individuals may only have a record of a single restrictive intervention or may be subject to repeated incidents of restrictive interventions over time. Each individual on the RIDS system has a unique client identifier. The RIDS database contains data drawn from over 150 government disability institutions across Victoria, Australia. All individuals included on the database from 1 July 2007 up till the end of 2012 were eligible for inclusion.
The VPCR (established 1961) is the state wide public mental health database in Victoria, Australia. It is a contacts-based database and documents when and why an individual comes into contact with public mental health services as well as a variety of other data including diagnostic categories, the number of contacts with services and dates and periods of admission and discharge. The VPCR records mental health diagnoses according to the International Classification for Mental Disorders version 9 and 10 (ICD 9, 10), based on thorough clinician assessments. The database does not capture contacts with primary care providers or private services so may underreport some high prevalence disorders such as anxiety, depression and substance use disorders.
The Law Enforcement Assistance Program (LEAP) database is a state wide reporting system maintained by Victoria Police that details whenever an individual has come into contact with police as a suspect, offender, victim, witness or person in need of assistance. It has been in its current format since October 1993. Incidents of offending and victimisation were extracted in raw form and, consistent with the extant literature, categorised as either: (1) violent, (2) sexual, or (3) non-violent non-sexual offences. Violent offences included common assault, murder, aggravated robbery, as well as any form of contact sexual offence; sexual offences involved indecent assault, rape and incest; and non-sexual non-violent offences included theft, property damage, substance misuse, threats, arson and non-contact stalking. The same three categories were used to classify both victimisation and offending histories. For the offending variables, the level of a criminal charge was selected. This point in criminal proceedings was selected in line with other recent research; a charge being indicative that there was a good deal of confidence that the alleged offence should be dealt with in a criminal court. Henceforth, for simplicity, the term offending is used to enable comparisons to be made.
Data linkage procedure
Participant information from all databases were compiled into a single file by matching participant-level details across each database using a master list (containing full name, date of birth and gender along with a unique study ID number). Deterministic and probabilistic matching algorithms were used to maximise potential matches between databases; where individual matches were found, all relevant contact-level data were extracted. Rates of contacts and counts of criminal charges and victimisation episodes were compared to those in a random community sample from a related study of 5000 males and females drawn from the Australian Electoral Role whose case ascertainment for mental health and police contact histories had been determined using an identical methodology and the same suite of databases [ 35 ]. Due to the nature of the data available for the community sample, the offending history variable used ‘conviction’ to classify an offence history outcome for the community sample.
Approach to analysis
Both individuals with ID and community comparison participants were compared to determine the prevalence rates of offending and victimisation in each group. The ID sample was split into two subgroups, differentiating: (1) those who had a primary diagnosis of ID and a secondary diagnosis of any mental illness (Comorbid ID group) from (2) those solely with an ID (ID only group). Prior to analysis the data were checked for missing values and a randomly selected 10 % of the ID cases ( n = 260) were recoded and crosschecked to check for spurious data entry errors.
Variables of interest were cross-tabulated and compared using Chi Squared tests of Association, substituting Fishers Exact test statistic where cell numbers in contingency tables fell below n = 5. Odds ratios and relative risk statistics were then calculated, along with 95 % confidence intervals applying Miettinen’s test-based approach [ 36 ]. All associations were considered statistically significant at an alpha level of .05. Effect sizes were calculated using Cohen’s d , with traditional cut-offs used to determine small, medium and large effects [ 37 ]. Data were also stratified according to gender, to ascertain whether the risks for particular offence or victimisation experiences differed between males and females.
General characteristics
The full sample comprised 2600 participants ( Males = 1684, 64.7 %, Females = 916, 34.2 %). The community comparison group included 4830 individuals ( M = 2392, 49.5 %, F = 2438, 50.5 %). The community group were significantly older than the ID group (ID = 35.71 (16.57), community = 39.12 (12.55), t =9.95 (7428), p < .0001) with males significantly younger in the ID group ( M = 34.12 (16.39), F = 38.66 (16.50), t = 7.13 (2911), p < .0001). There was no difference in age between males and females in the community group ( t = 0.826 (4477), p = 0.409). Of those diagnosed with an ID, over a quarter ( N = 709, 27.2 %) also met criteria for a comorbid mental illness and formed the Comorbid subgroup while the remaining ID sample ( N = 1891, 72.7 %) represented the ID only subgroup.
- Victimisation
The community group were significantly more likely to have an official history of victimisation compared to the full ID sample, with the risk of victimisation being two times higher. However, at the specific crime level the rate of victimisation increased significantly for the ID sample with the rate of violent victimisation two times higher, while sexual victimisation was nearly six times higher compared to the community; effect sizes were moderate to large (Table 1 ). Those with ID and a comorbid mental illness had the highest rates of victimisation with a three-fold increase for violent victimisation and a ten-fold increase for sexual victimisation compared to the community. In contrast, the ID only group had higher rates of victimisation relative to the community sample, however they were victimised at a rate less than the comorbid group. Across all victimisation episodes, the comorbid ID group had approximately double the risk of being victimised compared to the ID only group (Table 2 ).
Gender differences for victimisation
There were no significant differences in overall victimisation rates between males and females in the total ID group (χ 2 = 1.67, p = .1962). In the community sample there were similar proportions of males and females, however the overall risk for being victimised was 1.38 times higher for males than for females (RR = 1.38, 95 % CI = 1.29–1.49).
There were substantial differences between the ID sample and the community group; the risk of violent and sexual victimisation being three and five times (respectively) higher for females with ID compared to females in the community (RR = 3.07, 95 % CI = 2.44–3.86; RR = 5.05, 95 % CI = 3.45–7.39). Males with ID were violently victimised at a rate nearly two times that of males in the community (RR = 1.76, 95 % CI = 1.47–2.10) and sexually victimised at a rate in excess of 11 times higher (RR = 11.79, 95 % CI = 6.14–22.65). Females experienced violent victimisation at a significantly higher rate compared to males in the ID sample (RR = 3.45, 95 % CI = 2.68–4.45). Sex differences according to other victimisation types were not significantly different.
Criminal offending
Less than 10 % of the entire ID sample had a record of criminal charges according to the police LEAP database. There were no significant differences in the rates of offending between the ID sample overall (224/2600, 8.6 %) and the community sample (429/4830, 8.9 %). However, the ID sample violently offended at a rate three times higher than the community and sexually offended at a rate nearly eight times higher. By contrast, the rate of non-sexual non-violent offending was lower than that found in the community sample (Table 3 ).
The ID only group violently offended at a rate 1.6 times greater, and sexually offended at a rate 3.6 times that of the community sample. The most pronounced differences were again between the comorbid sample and the community sample with the comorbid sample offending at a rate 6.5 times greater than the community for violent crimes, with this rate increasing to 18.9 times higher for sexual offences. The increased rates of offending in the comorbid group were similar even when compared to the ID only group, with a four-fold increase in violent offending and a five-fold increase in sexual offending (Table 4 ).
Gender differences for offending
Among the Total ID sample, males were more likely to have a record of criminal offences compared to females (χ 2 = 17.53, p < .0001), with this result also reflected in the community sample with offence convictions (χ 2 = 196.43, p < .0001). Both males and females in the total ID group had significantly higher rates of criminality compared to males and females in the community group. The most pronounced difference was for females, with females with ID violently offending at a rate 11 times higher than females in the community (RR = 11.64, 95 % CI = 5.42–25.01). Males violently offended at a rate double that of males in the community (RR = 2.01, 95 % CI = 1.59–2.54) with the rate of sexual offending being nearly six times higher (RR = 5.84, 95 % CI = 3.50–9.74). Comparison of confidence intervals showed that the relative risk between the total ID group and the community was significantly higher for females than for males.
This study investigated victimisation and offending histories in a sample of people with intellectual disability and a community comparison sample using a case linkage design. The results indicated that, overall, people with intellectual disability were less likely to have an official history of victimisation and were no more or less likely to have a history of criminal offending than people without intellectual disability. Of note, however, the ID group were significantly more vulnerable to violent and sexual victimisation and offending compared to the community.
These findings suggest that members of the general community are more likely to have a police record as a victim of crime overall, with the vast majority of these crimes being non-violent and non-sexual in nature. This finding is consistent with some prior research [ 2 , 38 ], but contrary to previous theory and other research, which has suggested pronounced vulnerabilities for people with ID across all crime types [ 5 , 6 , 10 , 18 ]. Two explanations may account for this finding. First, individuals in the ID sample may have had less exposure to certain types of victimisation experiences due to the nature of their community and/or residential living circumstances. Alternatively, non-violent non sexual crimes may be under-reported by people with ID, who may not be aware of appropriate avenues for reporting, may be unable to recognise more ambiguous non-violent non-sexual crimes, may not be progressed to police services by carers/residential staff, or that they may fear reporting a person who they depend on [ 5 ].
Baladerian and others [ 23 ] noted that less than half of violent and sexual crimes against people with ID were reported to police, and of those reported, over half said nothing happened and less than 1 in 10 perpetrators were subsequently arrested. Participants in that study cited a lack of confidence in the criminal justice system, fear of retribution and poor knowledge of reporting avenues as key barriers to reporting. The potential for underreporting should be considered. Practically, efforts to support vulnerable populations report crime should be considered.
Despite the community having a higher rate of victimisation for crime overall, the current findings demonstrate that serious offences such as violent and sexual crimes are statistically more prevalent among people with ID, which supports the study hypotheses and previous research findings [ 11 , 13 , 19 – 22 ]. Roughly 1 in 6 people in the ID sample had reported violent victimisation to police, which was twice the rate of the community; furthermore 6 % had reported sexual victimisation, which was nearly six times higher than the community rate. These figures are concerning and provide robust epidemiological insight into the extent of an under researched problem and supports the conclusions of previous smaller scale studies which noted the heightened risk for violent and sexual crimes specifically [ 6 , 13 , 19 ].
Of particular interest to the current study was the influence of gender on victimisation. Findings indicate that males in the community group were more likely to have records of being victims than females, consistent with prior research [ 25 , 39 – 41 ]. Interestingly, this pattern of victimisation was not evident in the ID sample where there was no difference among victimisation types for gender, except for violent victimisation, which was significantly higher for females compared to males. This result is substantiated by the large difference in violent and sexual victimisation between the ID group and the community, for females more so than males, suggesting that females with ID are an especially vulnerable subgroup. What makes them specifically vulnerable to violent crime is unclear in the literature with only one report highlighting a possible gender difference [ 27 ]. The findings from the present study, coupled with the continued lack of consensus in the literature, should act as a catalyst to focus more on elucidating potential differences, and if so why these exist, as at present females with ID are particularly vulnerable to serious victimisation.
The rate of criminal offending, in this sample operationalized as criminal charges being laid, was entirely consistent with previous studies [ 14 , 33 ]. In this study, the overall rate of offending did not differ between the intellectual disabled and the community groups, with less than 10 % of both samples having an official record of offending. However, like victimisation, violent and sexual offending were statistically more common for people with ID, with offending six and a half times higher for violent crime and nearly 19 times higher for sexual crimes. This result can be interpreted using Routine Activities Theory [ 17 ] which postulates that a greater exposure to crime-inducing situations and personal reactions from the person with ID can make them more vulnerable to victimisation. This can also be applied to offending situations where an individual is similarly exposed to dangerous situations and there is still a potential for the individual to be a victim or an offender in an ambiguous and threatening situation. Empirical research corroborates this theoretical assumption and the current findings, which note a disproportionate number of people with ID in the criminal justice system and suggest their particular susceptibility to sexual offending [ 2 , 14 , 33 , 34 ]. In line with the Routine Activities Theory [ 17 ] the higher rates of offending may be related to the significant environmental and individual challenges faced by people with ID [ 2 ]. While offending may be more pronounced in people with ID, there is also a greater propensity for parental adversity, low socio economic status and mental illness. Future research should therefore seek to discern the relative importance of these variables to crime in ID populations.
Comorbid mental illness
The presence of a co-occurring mental illness significantly increased the likelihood of people with ID having both victimisation and offending histories. Mental illness has been associated with victimisation and offending in other vulnerable populations [ 42 – 45 ] with one study suggesting that mental illness may be linked with criminality in people with intellectual disability [ 46 ]. The current findings suggest that mental illness complicates the association between ID and victimisation and offending considerably (almost doubling the rates of both).
The presence of a mental illness may further limit the functionality of the individual with ID and may intensify their exposure to dangerous situations and reactions to potential perpetrators. The association between ID with comorbid mental illness and increased rates of victimisation and offending indicates that treating mental illness alongside managing deficits associated with ID could have beneficial effects for crime prevention and victimisation. However, while mental illness appears to be pivotal in influencing susceptibility to crime, there may be other contributing factors such as substance abuse, outside of the scope of the present study, which further complicates the relationship.
The present study complements the existing literature base providing a robust contemporary prevalence estimate for victimisation and offending among people with ID. It adds further weight to prior research findings regarding criminality among those with ID and proposes that people with ID are at a significant disadvantage and are over represented in crime figures. By illustrating the magnitude of the problem, using a robust epidemiological design, it is hoped that there will be greater research into why this problem exists and how this effect can be minimised.
Limitations
The findings in the current study may be limited by several factors that were inherent in the databases used. First, the Restrictive Interventions Database System (RIDS) may not be representative of all people with intellectual disabilities as individuals who are included on this reporting system have been subject to at least one restrictive intervention. This may have led to an over or underestimation of the true rates offending and victimisation reported here. While not available through the current dataset, taking into account a frequency criterion relating to the number of restrictive intervention episodes experienced by the person with ID as a potential confounding factor may help further develop our understanding of both risks and vulnerabilities to crime among people with ID who have more complex presentations or who present with more challenging behavioural management issues. Second, notwithstanding the challenges associated with diagnosing mental disorders among people with ID [ 47 , 48 ], rates of mental illness were estimated from a public mental health database which, as noted in the methodology, under-reports some of the more high prevalence disorders. That being said, a strength with the current methodology was that case ascertainment of mental disorder and police contact for individuals in the community comparison sample was identical, thereby leading to direct comparability and greater confidence in the magnitude and direction of associations reported here. Thirdly, the findings may underrepresent the extent of crime involvement in ID as victimisation and offending data were based on contact with the police, where official reports were made and subsequently recorded. From the literature, we know that individuals with ID typically under report crimes and may find it difficult to recognise these crimes. It is likely that the current estimate of crime is more conservative compared to true prevalence figures [ 23 ]. Further, it was not possible to statistically control for the potentially confounding effect of age as the ID and community databases were independent of each other. While this is unlikely to affect the direction of the associations with violent and sexual offending, the younger age of the ID sample may explain the finding pertaining to their lower risk of being the victim of other types of non-sexual non-violent crimes found with this sample, although the evidence remains inconclusive. Lastly, the community comparison sample operationalized offending at the level of conviction, while the ID sample used the level of criminal charge; this may mean that the statistical differences presented may represent an upper confidence limit for estimates increased risks of both perpetration and victimisation histories.
Directions for future research
Future research should seek to replicate and extend on current findings, which represent a preliminary yet robust insight into the vulnerabilities of those with ID. Of particular interest is future research is differences between specific age groups, mental illnesses and specific licit and illicit substances. Mental illness was a key factor in the association between intellectual disability and crime perpetration, the influence of specific mental illness on ID was not considered in the current study, as diagnoses are difficult to establish with comorbid ID. A future study with a greater focus on the robust assessment of mental illnesses and substance use could identify more specific disorders that are pertinent to both victimisation and offending; such information would be critical to informing both risk assessment and treatment planning.
Results of the current study provide robust prevalence estimates indicating that, statistically speaking, people with intellectual disability are at greater risk of experiencing violent and sexual victimisation and more likely to violently and sexually offend than non-disabled people living in the community. Future research should seek to elucidate why these differences can and do exist and should account for other contributing factors that may influence this relationship.
Abbreviations
intellectual disability
Law Enforcement Assistance Program
National Health & Medical Research Council
restrictive interventions data system
Victorian Psychiatric Case Register
Holland T, Clare IC, Mukhopadhyay T. Prevalence of criminal offending by men and women with intellectual disability and the characteristics of offenders. J Intellect Disabil Res. 2002;46:6–20.
Article PubMed Google Scholar
Simpson MK, Hogg J. Patterns of offending among people with intellectual disability: A systematic review. Part I: Methodology and prevalence data. J Intellect Disabil Res. 2001;45(5):384–96.
Article CAS PubMed Google Scholar
Hayes SC. People with an intellectual disability and the criminal justice: appearances before local courts. 1993. New South Wales Law Reform Commission.
Google Scholar
Winter N, Holland A, Collins S. Factors predisposing to suspected offending by adults with self-reported learning disabilities. Psychol Med. 1997;27(3):595–607.
Petersilia JR. Crime Victims with Developmental Disabilities A Review Essay. Crim Justice Behav. 2001;28(6):655–94.
Article Google Scholar
Wilson C, Brewer N. The incidence of criminal victimisation of individuals with an intellectual disability. Aust Psychol. 1992;27(2):114–7.
Evans DN. Perceptions of the Victimization of Individuals with Intellectual Disabilities. Adv Appl Socio. 2013;3(2):114–23.
Rand MR, Harrell E. Crime against people with disabilities, 2007. Washington, DC: Bureau of Justice Statistics; 2009.
Powers LE, Curry MA, McNeff E, Saxton M, Powers JL, Oschwald M. End the silence: A survey of abuse against men with disabilities. J Rehab. 2008;74(4):41–50.
Horner‐Johnson W, Drum CE. Prevalence of maltreatment of people with intellectual disabilities: A review of recently published research. Ment Retard and Dev Disabil Res Rev. 2006;12(1):57–69.
Verdugo MA, Bermejo BG, Fuertes J. The maltreatment of intellectually handicapped children and adolescents. Child Abuse Negl. 1995;19:205–15.
Brown BS, Courtless TF. The Mentally Retarded Offender, National Institute of Mental Health, Center for Studies of Crime and Delinquency. 1971.
Book Google Scholar
Sullivan PM, Knutson JF. Maltreatment and disabilities: A population-based epidemiological study. Child Abuse Negl. 2000;24(10):1257–73.
McBrien J, Hodgetts A, Gregory J. Offending and risky behaviour in community services for people with intellectual disabilities in one local authority. J Forens Psychi Psych. 2003;14(2):280–97.
American Psychiatric Association. Diagnostic and statistical manual of mental disorders (4th ed., text rev.). Washington, DC: American Psychiatric Association; 2000.
Nettelbeck T, Wilson C. Personal vulnerability to victimization of people with mental retardation. Trauma Viol Abuse. 2002;3(4):289–306.
Cohen LE, Felson M. Social change and crime rate trends: A routine activity approach. Am Sociol Rev. 1979;44:588–608.
Sobsey D. Patterns of sexual abuse and assault. J Sex Disabil. 1991;3:243–59.
Sobsey D. Sexual abuse of individuals with intellectual disability. In: Craft A, editor. Practice issues in sexuality and learning disabilities. New York: Routledge; 1994. p. 93–115.
McCarthy M, Thompson D. A prevalence study of sexual abuse of adults with intellectual disabilities referred for sex education. J Appl Res Intellect Disabil. 1997;10(2):105–24.
Balogh R, Bretherton K, Whibley S, Berney T, Graham S, Richold P, Firth H. Sexual abuse in children and adolescents with intellectual disability. J Intellect Disabil Res. 2001;45(3):194–201.
Furey EM. Sexual abuse of adults with mental retardation: Who and where. Ment Retard. 1994;32(3):173–80.
CAS PubMed Google Scholar
Baladerian NJ, Coleman TF, Stream J. Abuse of People with Disabilities: victims and their families speak out. 2013. Spectrum Institute Disability and Abuse Project, http://disability-abuse.com/survey/survey-report.pdf .
Reiter S, Bryen DN, Shachar I. Adolescents with intellectual disabilities as victims of abuse. J Intellect Disabil Res. 2007;11(4):371–87.
Lauritsen JL, Heimer K. The gender gap in violent victimisation 1973–2004. J Quant Crimin. 2008;24(2):125–47.
Harrell E, Truman J, Curto V, Thomas J, Quinn B, James D. Crime against persons with disabilities, 2009–2011. 2012. Statistical tables. US Department of Justice Office of Justice Programs Bureau of Justice Statistics.
Lindsay WR, Taylor JL, Sturmey P. Offenders with developmental disabilities. Washington: John Wiley & Sons; 2004.
Herrington V. Assessing the prevalence of intellectual disability among young male prisoners. J Intellect Disabil Res. 2009;53(5):397–410.
Holland S, Persson P. Intellectual disability in the Victorian prison system: Characteristics of prisoners with an intellectual disability released from prison in 2003–2006. Psychol Crime Law. 2011;17(1):25–41.
Webber LS, Lambrick F, Donley M, Buchholtz M, Chan JB, Carracher R, Patel G. Restraint and seclusion of people on compulsory treatment orders in Victoria, Australia in 2008–2009. Psychi Psychol Law. 2010;17(4):562–73.
Lindsay WR, Hastings RP, Beech AR. Forensic research in offenders with intellectual & developmental disabilities 1: prevalence and risk assessment. Psychol Crime Law. 2011;17(1):3–7.
Fazel S, Xenitidis K, Powell P. The prevalence of intellectual disabilities among 12,000 prisoners – a systematic review. Int J Law Psychiatry. 2011;31(4):369–74.
Vanny KA, Levy MH, Greenberg DM, Hayes SC. Mental illness and intellectual disability in magistrates courts in New South Wales. Australia J Intellect Disabil Res. 2009;53(3):289–97.
Hodgins S, Mednick SA, Brennan PA, Schulsinger F, Engberg M. Mental disorder and crime: evidence from a Danish birth cohort. Arch Gen Psychiatry. 1996;53(6):489–96.
Short T, Thomas S, Luebbers S, Ogloff JRP, Mullen P. Utilization of public mental health services in a random community sample. Aust NZ J Psychiat. 2010;44:475–81.
Kirkwood B. Essentials of Medical Statistics. Oxford: Blackwell Science; 2001.
Cohen J. A Power Primer. Psychol Bull. 1992;112:155–9.
McCleary R, Wiebe DJ. Measuring the victimization risk of the developmentally disabled: Methodological problems and solutions. Unpublished manuscript. Washington DC: National Research Council; 1999. Cited in Petersilia JR: Crime victims with Developmental Disability: A Review Essay. Crim Justice & Beh 2001, 28(6), 655–694.
Lewin B. Who cares about disabled victims of crime? Barriers and facilitators for redress. J Pol Pract Intellect Disabil. 2007;4(3):170–6.
Cooper A, Smith EL. Homicide Trends in the United States, 1980–2008. 2011. U.S Department of Justice.
Vaillancourt R. Gender Differences in Police-reported Violent Crime in Canada, 2008. 2010. Canadian Centre for Justice Statistics.
Arseneault L, Moffitt TE, Caspi A, Taylor PJ, Silva PA. Mental disorders and violence in a total birth cohort: results from the Dunedin Study. Arch Gen Psychiatry. 2000;57:979–86.
Steadman HJ, Mulvey EP, Monahan J, Robbins PC, Appelbaum PS. Violence by people discharged from acute psychiatric inpatient facilities and by others in the same neighborhoods. Arch Gen Psychiatry. 1998;55:393–401.
Swanson JW, Holzer CE, Ganju VK, Jono RT. Violence and psychiatric disorder in the community: evidence from the Epidemiologic Catchment Area surveys. Hosp Comm Psychiat. 1990;41:761–70.
CAS Google Scholar
Tiihonen J, Isohanni M, Rasanen P, Koiranen M, Moring J. Specific major mental disorders and criminality: a 26-year prospective study of the 1966 northern Finland birth cohort. Am J Psychiatry. 1997;154:840–5.
Hodgins S. Mental disorder, intellectual deficiency, and crime: evidence from a birth cohort. Arch Gen Psychiatry. 1992;49(6):476–83.
Cooper SA, Smiley E, Morrison J, Williamson A, Allan L. Mental ill-health in adults with intellectual disabilities: prevalence and associated factors. Br J Psychiatry. 2007;190(1):27–35.
Deb S, Thomas M, Bright C. Mental disorder in adults with intellectual disability. 1: Prevalence of functional psychiatric illness among a community‐based population aged between 16 and 64 years. J Intellect Disabil Res. 2001;45(6):495–505.
Health N, Council MR. National Statement on Ethical Conduct in Human Research 2007 (Updated May 2015). NHMRC: Commonwealth of Australia; 2007. https://www.nhmrc.gov.au/_files_nhmrc/publications/attachments/e72_national_statement_may_2015_150514_a.pdf .
Download references
Acknowledgements
We would like to thank Leanne Sargent and Dr David Ballek at Victoria Police, and Dr Jeffrey Chan, Dr Lynne Webber and Dr Frank Lambrick at the Office of the Senior Practitioner.
The project was funded as part of an Australian Research Council Linkage Grant with the Victorian Office of the Senior Practitioner and Victoria Police. Representatives from the Office of the Senior Practitioner and Victoria Police helped develop the design and scope of the study and provided dedicated personnel to collate and extract the contact-based data required for the linkage methodology. Representatives from these two organisations also received copies of the draft manuscript for their information and were invited to respond with any comments and/or suggestions.
Availability of data and materials
Due to the sensitive nature of the contact-level data used from multiple agencies, and constraints of the ethical approvals, the linked data are not publically available.
Authors’ contributions
BF coded, analysed and interpreted the data and contributed to the drafting of the manuscript. ST was involved in the conception, design, analysis and interpretation of the research findings and contributed to the drafting of the manuscript. MD was involved in the conception of the study, interpretation of the research findings and contributed to the drafting of the manuscript. JO was involved in the conception of the study, interpretation of the research findings and contributed to the drafting of the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent to publish
Not applicable.
Ethics and consent to participate
The study design utilised existing contact-based data collected routinely by the Department of Human Services and Victoria Police. The data linkage methodology is supported through guidelines provided in the National Statement on Ethical Conduct in Human Research [ 49 ] relating to the use of databanks. Of note, Section 3.2.4 of the document states that ‘approval may be given to the use of identifiable data to ensure that the linkage is accurate, even if consent has not been given for the use of identifiable data in research’ (pg. 28). The project was approved by University of Wollongong Human Research Ethics Committee and Victoria Police Human Research Ethics Committee.
Author information
Authors and affiliations.
Faculty of Social Sciences, University of Wollongong, New South Wales, 2522, Australia
Billy C. Fogden & Stuart D. M. Thomas
School of Global, Urban and Social Studies, RMIT University, Building 37, Level 4, Swanston Street, Melbourne, Victoria, 3001, Australia
Stuart D. M. Thomas
Centre for Forensic Behavioural Science, Swinburne University of Technology, Melbourne, Australia
Michael Daffern & James R. P. Ogloff
Southern Clinical School, Monash University, Melbourne, Australia
Stuart D. M. Thomas & James R. P. Ogloff
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Stuart D. M. Thomas .
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License ( http://creativecommons.org/licenses/by/4.0/ ), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated.
Reprints and permissions
About this article
Cite this article.
Fogden, B.C., Thomas, S.D.M., Daffern, M. et al. Crime and victimisation in people with intellectual disability: a case linkage study. BMC Psychiatry 16 , 170 (2016). https://doi.org/10.1186/s12888-016-0869-7
Download citation
Received : 21 December 2015
Accepted : 16 May 2016
Published : 28 May 2016
DOI : https://doi.org/10.1186/s12888-016-0869-7
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Intellectual disability
- Mental disorder
BMC Psychiatry
ISSN: 1471-244X
- Submission enquiries: [email protected]
- General enquiries: [email protected]
ADNP dysregulates methylation and mitochondrial gene expression in the cerebellum of a Helsmoortel–Van der Aa syndrome autopsy case
- Open access
- Published: 18 April 2024
- Volume 12 , article number 62 , ( 2024 )
Cite this article
You have full access to this open access article

- Claudio D’Incal ORCID: orcid.org/0000-0003-1888-9066 1 , 2 ,
- Anke Van Dijck 1 , 3 ,
- Joe Ibrahim 1 ,
- Kevin De Man 1 , 2 ,
- Lina Bastini 2 ,
- Anthony Konings 2 ,
- Ellen Elinck 1 ,
- lllana Gozes 4 ,
- Zlatko Marusic 5 ,
- Mirna Anicic 6 ,
- Jurica Vukovic 6 ,
- Nathalie Van der Aa 1 ,
- Ligia Mateiu 1 ,
- Wim Vanden Berghe 2 &
- R. Frank Kooy 1
474 Accesses
Explore all metrics
Helsmoortel–Van der Aa syndrome is a neurodevelopmental disorder in which patients present with autism, intellectual disability, and frequent extra-neurological features such as feeding and gastrointestinal problems, visual impairments, and cardiac abnormalities. All patients exhibit heterozygous de novo nonsense or frameshift stop mutations in the Activity-Dependent Neuroprotective Protein ( ADNP ) gene, accounting for a prevalence of 0.2% of all autism cases worldwide. ADNP fulfills an essential chromatin remodeling function during brain development. In this study, we investigated the cerebellum of a died 6-year-old male patient with the c.1676dupA/p.His559Glnfs*3 ADNP mutation.
The clinical presentation of the patient was representative of the Helsmoortel–Van der Aa syndrome. During his lifespan, he underwent two liver transplantations after which the child died because of multiple organ failure. An autopsy was performed, and various tissue samples were taken for further analysis. We performed a molecular characterization of the cerebellum, a brain region involved in motor coordination, known for its highest ADNP expression and compared it to an age-matched control subject. Importantly, epigenome-wide analysis of the ADNP cerebellum identified CpG methylation differences and expression of multiple pathways causing neurodevelopmental delay. Interestingly, transcription factor motif enrichment analysis of differentially methylated genes showed that the ADNP binding motif was the most significantly enriched. RNA sequencing of the autopsy brain further identified downregulation of the WNT signaling pathway and autophagy defects as possible causes of neurodevelopmental delay. Ultimately, label-free quantification mass spectrometry identified differentially expressed proteins involved in mitochondrial stress and sirtuin signaling pathways amongst others. Protein–protein interaction analysis further revealed a network including chromatin remodelers (ADNP, SMARCC2, HDAC2 and YY1), autophagy-related proteins (LAMP1, BECN1 and LC3) as well as a key histone deacetylating enzyme SIRT1, involved in mitochondrial energy metabolism. The protein interaction of ADNP with SIRT1 was further biochemically validated through the microtubule-end binding proteins EB1/EB3 by direct co-immunoprecipitation in mouse cerebellum, suggesting important mito-epigenetic crosstalk between chromatin remodeling and mitochondrial energy metabolism linked to autophagy stress responses. This is further supported by mitochondrial activity assays and stainings in patient-derived fibroblasts which suggest mitochondrial dysfunctions in the ADNP deficient human brain.
This study forms the baseline clinical and molecular characterization of an ADNP autopsy cerebellum, providing novel insights in the disease mechanisms of the Helsmoortel–Van der Aa syndrome. By combining multi-omic and biochemical approaches, we identified a novel SIRT1-EB1/EB3-ADNP protein complex which may contribute to autophagic flux alterations and impaired mitochondrial metabolism in the Helsmoortel–Van der Aa syndrome and holds promise as a new therapeutic target.
Graphical abstract
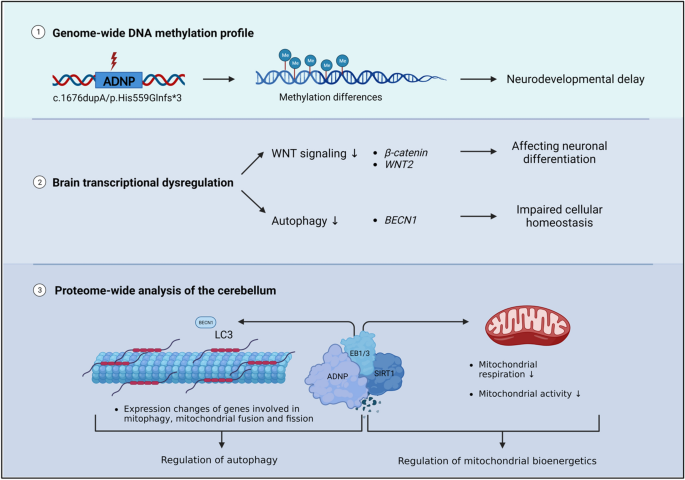
The ADNP patient mutation affects genome-wide methylation and leads to neurodevelopmental abnormalities.
The ADNP brain transcriptome reveals impaired neuronal differentiation and cellular homeostasis by aberrant signaling of the WNT pathway and autophagy process.
ADNP forms a complex with SIRT1 through the microtubule end-binding proteins EB1 and EB3.
Mitochondrial gene expression is impaired in the ADNP brain and patient-derived cellular models.
Similar content being viewed by others

Rare genetic variation in fibronectin 1 (FN1) protects against APOEε4 in Alzheimer’s disease

Abundant transcriptomic alterations in the human cerebellum of patients with a C9orf72 repeat expansion

Spinocerebellar ataxia: an update
Introduction.
The development of whole exome sequencing (WES) has substantially increased our insights in the genetic causes of neurodevelopmental disorders by detection of de novo mutations by comparing the exome of the proband to that of its parents [ 1 ]. Using this method, mutations in the Activity Dependent-Neuroprotective Homeobox Protein ( ADNP ) gene have been discovered, contributing to a neurogenetic syndrome called Helsmoortel–Van der Aa syndrome ( OMIM 615873 ), with a prevalence of 0.2% of global autism cases [ 2 , 3 ]. Patients show a clinical presentation of mild to severe intellectual disability (ID), autism spectrum disorder (ASD), global developmental delay (GDD), motor and speech delay, behavior abnormalities and deficiencies in several organ systems such as gastrointestinal problems [ 4 ]. In recent screening studies, ADNP appears one of the most frequently mutated genes with a hundred percent disease penetrance [ 5 , 6 ].
While massive screening studies now have cumulated in the discovery of over a thousand genes that are involved in ID and/or ASD, our molecular and functional understanding of the pathophysiology of these genes is lagging far behind. For instance, despite a wealth of information, many biochemical aspects of the function of ADNP in the brain remain unknown [ 7 ]. Spanning a genomic length of almost 40 kb, the longest transcript of the ADNP gene contains six exons of which only the last three translate to the actual protein [ 8 ]. Functional domains of ADNP include nine zinc fingers, a bipartite nuclear localization signal (NLS), a homeobox domain with ARKS motif, a heterochromatin protein 1 (HP1)-interacting PxVxL motif, and the neuroprotective octapeptide sequence NAP VSIPQ (NAP) [ 8 , 9 , 10 , 11 , 12 ]. In the nucleus, ADNP plays a role in chromatin remodeling: it binds directly to other chromatin remodelers, including the BAF complex members BRG1, ARID1A, and SMARCC2 by its C-terminal tail as demonstrated in a HEK293 human embryonic kidney cell line [ 9 ], and to CHD4 by its N-terminus as well as HP1β by its C-terminus in the repressive ChAHP complex discovered in murine embryonic stem cells, where it competes with CTCF for a common set of binding motifs [ 13 ]. Besides, a stable triplex of ADNP, BRG1 and CHD4 was also reported in murine stem cells [ 14 ], while POGZ and HP1γ form a nuclear complex with ADNP in the embryonic mouse cortex [ 15 ]. Most recently, ADNP was predicted to interact with the WRD5-SIRT1-BRG1-HDAC2 including YY1 complex [ 16 ]. Although the involvement of ADNP in chromatin remodeling functions has been firmly established, the role of these protein complexes in the human brain remains to be determined. In terms of function, ADNP is involved in neuronal tube closure and brain development, controlling expression of hundreds if not thousands of genes [ 17 ].
The chromatin function of ADNP is reflected by specific, aberrant methylation patterns in the blood of patients. In fact and almost unique to ADNP, two partially opposing methylation patterns have been described, depending on the location of the mutation. Whereas mutations located at the 3′-end and 5′-end of the ADNP gene (outside of nucleotides c.2000–2340) represent a Class I episignature with a pattern of overall hypomethylated CpGs, mutations in the central region (within nucleotides 2000–2340) of the gene show rather CpG hypermethylation [ 18 , 19 ]. Interestingly, the hypermethylated region, encompassing the recurrent p.Tyr719* ADNP mutation, is associated with a more severe clinical presentation [ 4 , 20 ]. Cytoplasmic roles for ADNP have also been suggested e.g., involvement in autophagy by binding LC3 [ 21 ], and interactions with the cytoskeleton via the microtubule end-binding proteins (EB1/EB3) [ 22 , 23 ], with Adnp deficiency resulting in impaired axonal transport and impaired dendritic spines [ 23 , 24 , 25 , 26 ]. Additionally, ADNP interacts with other cytoskeletal proteins such as SHANK3 and actin as well as with the armadillo sequence of beta-catenin, important for WNT signaling [ 14 , 27 , 28 ].
However, none of the above-mentioned studies have been performed in disease-relevant tissue. Instead, immortalized human cell lines, murine tissues, embryonic stem cells or other model systems have been investigated. Here, we present a unique case study on autopsy material of a six-year-old child with the heterozygous c.1676dupA/p.His559Glnfs*3 de novo ADNP mutation . By combining in-depth epigenetic, transcriptomic, and proteomic studies in the cerebellum of this post-mortem ADNP subject, we were able to confirm the involvement of pathways such as the WNT-signaling in the Helsmoortel–Van der Aa syndrome as well as to demonstrate ADNP involvement in autophagy and mitochondrial (dys)function(s).
Materials and methods
Post-mortem tissues and subjects.
Clinical information of a nine-year-old female patient was obtained under informed written consent from the Institute Born-Bunge vzw IBB NeuroBioBank of the University of Antwerp and transferred with written informed consent under HMTA20210040 after approval of the Ethics Committee of the Antwerp University Hospital/University of Antwerp. The female subject, used as a control in this study, showed symptoms analogous to a sporadic form of Rett syndrome and died following obstructive apnea. Twelve hours after death, cerebellar tissue was collected during the autopsy and frozen in liquid nitrogen or fixed in formaldehyde. Frozen section, paraffin sections, and celloidin embeddings were extensively investigated by an expert pathologist, resulting in no morphological abnormalities in all brain regions, except some fibrillar gliosis in the hippocampus. Comparisons to other sections of an age-matched control showed similar cytology and no neuronal loss. The substantia nigra contained normal amounts of melanin granules and the cerebellum contained no loss of Purkinje cells. Clinical information of a dead six-year-old male patient with the heterozygous c.1676dupA/p.His559Glnfs*3 ADNP mutation was received under informed consent under B300201627322 and approved by the Ethics Committee of the Antwerp University Hospital. The patient died because of multiple organ failure. Cerebellar tissue was collected during autopsy following a 35-h post-mortem interval, subsequently frozen in liquid nitrogen or fixed in formaldehyde. Clinical evaluation was performed by at least one expert clinical geneticist. The ADNP mutation was confirmed by Sanger sequencing using the forward primer 3′-TGATGTGCAAGTGCATCAGA-5′ and reverse primer 3′-TGTGCACTTCGAAAAAGAACAT-5′. Conservation of the amino acids changed by the ADNP mutation was verified using ClustalW.
Plasmid constructs and site-directed mutagenesis
The pCMV3 expression vector encoding human wild-type ADNP fused to either an N-terminal GFPSpark® or N-DYKDDDDK (Flag®) tag was purchased from Sino Biological ( HG11106-ANG ; HG11106-NF ). The c.1676duplA mutation was introduced in the N-DYKDDDDK (Flag®) ADNP expression vector by PCR mutagenesis using the Q5® Site-Directed mutagenesis kit (New England Biolabs; E0554S ) according to manufacturer’s protocol. Mutagenesis primers were designed using the NEBaseChanger Tool ( http://nebasechanger.neb.com/ ). The mutation was inserted with the forward primer: 5′-ACACTAACATCCATCTCCTG-3’ and the reverse primer: 5′-TGACTACCCTGCTGCAAT-3′ by thermocycling with an annealing temperature of 60 °C. DNA was purified from transformed high-efficiency NEB 5-alpha competent E. coli cells using the NucleoSpin Plasmid EasyPure Mini kit (Macherey Nagel; 740727.50 ) according to the manual. The mutation was confirmed by Sanger sequencing.
Cell lines and culture conditions
HEK293T cells (ATCC; CRL-3216™ ) were cultured at low passage number in DMEM (Gibco™; 11965092 ), supplemented with 10% fetal bovine serum (Gibco™; 26140079 ) and 1% penicillin/streptomycin (Gibco™; 15070063 ). Age- and sex matched Epstein-Barr virus transformed lymphoblastoid cell lines (LCLs) of healthy subjects (n = 4) and patients with different ADNP mutations (n = 6) (Additional file 1 : Table S1) were cultured in RPMI (Gibco™; A1049101), supplemented with 15% fetal bovine serum (Gibco™; 26140079), 1% penicillin/streptomycin (Gibco™; 15070063), 1% sodium pyruvate (Gibco™; 11360070), and 1% GlutaMAX (Gibco™; 35050061). Age- and sex matched skin fibroblasts of two unrelated asymptomatic subjects and two patients with different ADNP mutations (n = 2) (Additional file 1 : Table S1) were cultured in RPMI (Gibco™; A1049101), supplemented with 15% fetal bovine serum (Gibco™; 26140079), 1% penicillin/streptomycin (Gibco™; 15070063), 1% sodium pyruvate (Gibco™; 11360070), and 1% GlutaMAX (Gibco™; 35050061). Human primary cell lines were obtained from consenting individuals, guardians, tending clinicians, or parents. All procedures were carried out following the guidelines and regulations of the University of Antwerp/University Hospital of Antwerp (UZA) and approved by the Ethics Committee of the Antwerp University Hospital. All cell lines were cultured in a humidified incubator at 37%O 2 /5%CO 2 .
AlfaFold 3D-structural protein modeling
The predicted 3D-structure of human wild-type ADNP (Uniprot; Q9H2P0) was acquired using the AlfaFold Protein Structure Database ( https://alphafold.ebi.ac.uk/ ). The p.His559Glnfs*3 mutant was queried in the amino acid sequence of wild-type ADNP. Wild-type and mutant ADNP proteins were modeled using AlfaFold2 with ColabFold ( https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb ), an online integrating AlfaFold2 pipeline for protein structure modeling combined with many-against-many sequence searching (MMSeqs2), and HHSearch. ChimeraX (UCSF, version 1.5) was used for visualization and annotation of the structural ADNP protein domains using the generated PBD output file as input.
Cellular ADNP transfection system
HEK293T were transiently transfected with 5 µg of human expression vectors: (1) wild-type ADNP with an N-terminal GFPSpark®-tag, (2) wild-type ADNP with an N-terminal DYKDDDDK (Flag®)-tag, or (3) mutant c.1676dupA ADNP fused to an N-terminal DYKDDDDK (Flag®)-tag using Lipofectamine™ 3000 Transfection Reagent (Invitrogen; L3000008 ) in accordance with the manufacturer’s protocol. Co-transfections were performed with equal amounts of both wild-type and mutant ADNP expression vectors. Transfection efficiency was about 70% in line with the manufacturer’s tested performance. Cells were harvested after 24-h for subcellular protein fractionation followed by western blotting.
ADNP expression analysis: total protein extraction and subcellular protein fractionation
After transient transfection of the ADNP expression vector of interest, HEK293T cells were detached with TrypLE™ Express Enzyme (1X), phenol red (Gibco™; 12605028 ), subsequently washed with ice-cold DPBS (Gibco™; 14040133 ). Cerebellar tissue obtained from the post-mortem control subject and died ADNP patient was homogenized with the TissueRuptor II (Qiagen; 9002755 ) with mixing at the lowest speed. For total protein extraction, cells and tissue were lysed in ice cold RIPA buffer (150 mM NaCl, 50 mM Tris, 0.5% sodium deoxycholate, 1% NP-40 and 2% sodium dodecyl sulfate), supplemented with the cOmplete™, Mini, EDTA-free Protease Inhibitor Cocktail (Roche; 04693159001 ) together with PhosSTOP™ phosphatase inhibitor (Roche; 4906845001). Lysis occurred for 15 min at 4 °C with agitation and cell debris was removed by centrifuging 15 min at maximal speed in a precooled centrifuge. For subcellular fractionation of transfected HEK293T cells, a final amount of 10 × 10 6 cells was lysed and gradually separated in cytoplasmic, membrane, nuclear soluble, chromatin-bound, and cytoskeletal protein extracts using the Subcellular Protein Fractionation Kit for Cultured Cells (Thermo Scientific™; 78840 ) following the manufacturer’s instructions. The protein concentration was estimated with the Pierce™ BCA Protein Assay Kit (Thermo Scientific™; 23225 ). ADNP expression was investigated in the cytoplasmic, chromatin-bound, and cytoskeletal protein fractions.
Immunoblotting
A total amount of 20 μg protein lysate was reduced with NuPAGE™ Sample Reducing Agent (Invitrogen; NP0009 ) in NuPAGE™ LDS Sample Buffer (Invitrogen; NP0007 ). Samples were heated for 10 min at 70 °C and subsequently loaded for separation using a Bolt™ 4 to 12%, Bis–Tris, 1.0 mm, Mini Protein Gels (Invitrogen; NW04120BOX ) using Bolt™ MOPS SDS Running Buffer (Invitrogen; B0001 ) at 120 V. The Precision Plus Protein™ All Blue Prestained Protein Standard (Biorad; #1610373 ) was used for estimation of the molecular weight in all experiments. After separation, proteins were transblotted onto Amersham™ Protran® Premium nitrocellulose membranes (Cytiva; GE10600008 ) using a Mini Trans-Blot® cell (Biorad; 1703930 ) with a transfer buffer containing 25 mM Tris, 192 mM glycine and 20% methanol (pH 8.3). Successful protein transfer was checked with a Ponceau S solution (Sigma Aldrich; P7170 ). Nitrocellulose membranes were blocked with either 5% blocking-grade non-fat dry milk (NFDM) (Carl Roth; T145.4 ) or 5% bovine serum albumin (BSA) (Carl Roth; CP84.1 ) dissolved in tris-buffered saline (TBST) for one hour at room temperature with agitation. Primary antibodies (Additional file 1 : Table S2) were tested and optimized to raise the least amount of background signals. Signal amplification was achieved by incubation with an appropriate HRP-conjugated immunoglobulins (Agilent) in a 1:2000 dilution in either 5% blocking-grade non-fat dry milk/TBST or 5% BSA/TBST solution. The signal was detected using the Pierce™ ECL Western Blotting Substrate (Thermo Scientific™; 32106 ). The West Femto Maximum Sensitivity Substrate (Thermo Scientific™; 34095 ) was used For ADNP detection specifically [ 29 ]. Image acquisition was executed with the Amersham™ Imager 680 (Cytiva). Monoclonal GAPDH (Cell signaling technology; 4317 ), Histone H3 (Abcam; 10799 ), and β-actin (Sigma-Aldrich; A5441 ) (Table S3) were used as loading controls for all the experiments. Image Quantification was performed using ImageJ software. Graphical representation was performed in GraphPad Prism version 9.3.1 using an unpaired student T-test assuming equal variances and normal distribution. Full western blot images are show as supplementary materials (Additional file 2 : Data S11).
Human methylation EPIC BeadChip array and data processing
Total DNA was isolated from the cerebellar tissue of the post-mortem control subject and patient cerebellum (n = 1) using the DNeasy Blood and Tissue Kit (Qiagen; 69504 ) according to the manufacturer’s instructions. Subsequently, bisulfite conversion of 250 ng isolated DNA was performed using the EZ DNA Methylation Kit (Zymo Research, D5001). To confirm successful bisulfite conversion, a methylation-conserved fragment of the human SALL3 gene was amplified using the following primers: 5′-GCGCGAGTCGAAGTAGGGC-3′ as forward primer and 5′-ACCCAACGATACCTAATAATAAAACC-3 as reverse primer with the PyroMark PCR kit (Qiagen; 978703 ). Amplified products were separated on a 1.5% agarose gel stained with GelRed® Nucleic Acid Gel Stain (Biotium; 41002 ). The TrackIt™ 100 bp DNA Ladder (Invitrogen; 10488-058 ) was used as a reference marker. Bisulfite-converted samples were hybridized on the Infinium Human Methylation EPIC BeadChip (Illumina; 20020531 ) as described in the manufacturer’s protocol. EPIC chips will be analyzed using the Illumina Hi-Scan system, a platform integrating more than 850,000 methylation sites quantitatively across the genome at single-nucleotide resolution. Raw intensity files were first quality checked and processed using the minfi package (v 1.38.0) [ 30 ]. Signal intensities were normalized using quantile normalization and beta values were calculated. Probes with a detection p -value higher than 0.01 were excluded. Non-CpG probes, probes with known single nucleotide polymorphisms (SNPs), multihit probes and probes on the X-/Y-chromosomes were filtered out. Probe annotation was carried out using the Illumina Infinium MethylationEPIC v1.0 B5 manifest file. All annotations (i.e., CpG islands, shelve, and shore regions) are reported based on the GRCh37/hg19 human genome build. We calculated the difference in methylation of the signals acquired in patient versus control subject with a focus on CpG probes showing over 20% methylation, i.e., hypomethylation (Δβ-values < −0.2) and hypermethylation (Δβ-value > 0.2). We determined gene ontology enrichment using the Metascape webtool [ 31 ]. Protein–protein network interactions of ADNP with the identified hypermethylated and hypomethylated genes were predicted using the STRING database version 12.0. The iRegulon plugin in Cytoscape was used to detect the transcription factors, their targets, and the motifs/tracks associated with co-expression of the hypomethylated and hypermethylated genes.
Targeted pyrosequencing analysis
Biologically-relevant genes exhibiting a methylation difference of Δβ > 0.2 (hypermethylation) and Δβ < −0.2 (hypomethylation) between patient and control cerebellum were selected for pyrosequencing validation. Briefly, the required primers (i.e., forward, reverse, and sequencing primers) were designed using the PyroMaker Assay Design 2.0 software (Qiagen) according to the manufacturer’s instructions (Additional file 1 : Table S3). Bisulfite-converted DNA fragments were PCR amplified using the PyroMark PCR kit (Qiagen; 978703 ). Successful PCR amplification was assessed by tris-boric acid-EDTA (TBE) electrophoresis at 1.5% agarose gel, after which the PyroMark Q24 Instrument (Qiagen) was used to perform pyrosequencing. Biotinylated PCR products were immobilized on streptavidin-coated Sepharose beads (GE Healthcare; 17511301 ), captured by the PyroMark vacuum Q24 workstation, washed and denatured. Single-stranded PCR products were subsequently released into a 24-well plate and annealed to the sequencing primer for 5 min at 80 °C. After completion of the pyrosequencing run, results were analyzed using the PyroMark Q24 software (Qiagen). Graphical representation was performed with GraphPad Prism version 9.3.1.
Total RNA extraction and sequencing of post-mortem brains and ADNP lymphoblastoid cell lines
Total RNA was extracted from the cerebellum of the control subject and the patient with the c.1676duplA/p.His559Gln*3 ADNP mutation (n = 1) as well as from control and patient LCLs with different ADNP mutations (n = 4 controls, n = 6 patients) using the RNeasy Mini Kit (Qiagen; 74106 ) according to the manufacturer’s protocol. RNA concentration was determined with the Qubit™ RNA Broad Range Assay Kit (Invitrogen™; Q10211 ) and the 260/280 ratio, indicative of RNA purity, was checked using NanoDrop™ 2000/2000c Spectrophotometer (Thermo Scientific™; ND-2000 ). RNA integrity was verified with Agilent RNA Screentape Assay on the 2200 TapeStation instrument (Agilent; G2964AA ). Samples with the highest RIN score (RIN > 6.5) were selected and sent to Novogene for RNA sequencing (RNAseq) (Additional file 1 : Table S1). All sequencing data was mapped to the human annotated genome GRCh38.p13 (Ensembl v106) with STAR, after adapter removal and reads cleaning with trimmomatic. Gene expression quantification was performed with featureCounts (subread package). We calculated gene expression differences in our post-mortem brains using NOISeq (R package), a non-parametric method for one-versus-one cases that reports the log2-ratio of the two conditions (M) and the value of the difference between conditions (D). A gene is considered to be differentially expressed if its corresponding M and D values are likely to be higher than in noise. A similar analysis was performed for the functional enrichment exploration for the up- and downregulated genes found by NOISeq at q > 0.95. Differential gene expression analysis for the LCL samples was performed with the DESeq2, an R package. The genes having a BH-adjusted FDR, p value < 0.05, and an absolute value of log2FC > = 0.5 were considered biologically relevant and further analyzed for functional enrichment (clusterProfiler R package with fGSEA function for the gene set enrichment analysis and enrichGO for overrepresentation analysis in GO ontologies and KEGG pathways). Additional data visualization was supported by BigOmics, a user-friendly and interactive cloud computing based bioinformatics platform for the in-depth analysis, visualization, and interpretation of transcriptomics data [ 32 ]. Ultimately, we performed a meta-analysis of the differentially expressed genes identified in the LCLs and post-mortem brains based on gene ID intersection and looked for conserved ADNP-relevant genes beyond their tissue-specific expression (brains versus LCLs).
RT-PCR gene expression analysis
RT-PCR was used to confirm a selection of genes from the RNA sequencing experiment (LCLs, post-mortem brains and common genes between data sets) by converting 1 µg of total extracted RNA to cDNA using the SuperScript™ III Reverse Transcriptase kit (Invitrogen™; 18080093 ). Primer efficiencies were optimized using a standard dilution curve method on pooled cDNA samples from controls and patients per dataset (90% > E > 110%). RT-PCR was performed in triplicate using the CFX384 Touch Real-Time PCR Detection System (BioRad; 1855484 ) with primers listed in Additional file 1 : Table S4 using the Takyon™ No ROX SYBR 2X MasterMix (Eurogentec; UF-NSMT-B0701 ). Reference gene stability was assessed using the geNorm method in qbase + (Biogazelle), after which were selected for normalization. Data analysis was performed in qbase + (Biogazelle) with a maximum deviation of 0.5 per triplicate using the stable housekeeping genes ACTB , B2M , and UBC . Statistical analysis was performed in GraphPad Prism 9.3.1 using an unpaired student T-test assuming unequal variances (post-mortem brains) and a Mann Witney U-test for unpaired measure (LCLs).
Label-free quantification (LFQ) mass spectrometry
Cerebellar tissue obtained from the post-mortem control subject and patient was homogenized with the TissueRuptor II (Qiagen; 9002755 ) with mixing at the lowest speed. Tissues were lysed and homogenized in ice cold RIPA buffer (150 mM NaCl, 50 mM Tris, 0.5% sodium deoxycholate, 1% NP-40 and 2% sodium dodecyl sulfate), supplemented with the cOmplete™, Mini, EDTA-free Protease Inhibitor Cocktail (Roche; 04693159001) together with PhosSTOP™ phosphatase inhibitor (Roche; 4906845001 ). Lysis occurred for one hour at 4 °C with agitation and cell debris was removed by centrifuging 30 min at maximal speed in a precooled centrifuge. The protein concentration was estimated with the Pierce™ BCA Protein Assay Kit (ThermoScientific™; 23225 ). Protein reduction, alkylation and digestion were performed with the ProteoSpin™ On-Column Proteolytic Digestion Kit (Norgen; 17500 ) according to manufacturer’s protocol. A nano-liquid chromatography (LC) column (Dionex ULTIMATE 3000) coupled online to a Q Exactive™ Plus Hybrid Quadrupole-Orbitrap™ Mass Spectrometer (Thermo Scientific™) was used for the MS analysis. Peptides were loaded for five technical replicates onto a 75 μm × 150 mm, 2 μm fused silica C18 capillary column, and mobile phase elution was performed using buffer A (0.1% formic acid in Milli-Q water) and buffer B (0.1% formic acid in 80% acetonitrile/Milli-Q water). The peptides were eluted using a gradient from 5% buffer B to 95% buffer B over 120 min at a flow rate of 0.3 μL/min. The LC eluent was directed to an ESI source for Orbitrap analysis. The MS was set to perform data dependent acquisition in the positive ion mode for a selected mass range of 375–2000 m/z for quantitative expression difference at the MS1 (140,000 resolution) level followed by peptide backbone fragmentation with normalized collision energy of 28 eV, and identification at the MS2 level (17,500 resolution). The *.RAW files were exported and processed in PEAKS AB 2.0 (Bioinformatics Solutions Inc.). The files were searched using target-decoy matching using the human UniProt database, with the false discovery rate set at 1%. Trypsin was indicated as the enzyme and up to two miscleavages were allowed. Carbamidomethylation was set as a fixed modification. Label-Free Quantification (LFQ) and Match Between Runs were used using default settings. PEAKS intensities were uploaded in MetaboAnalyst5.0, subsequently quantile normalized, log-transformed and autoscaled. An unpaired student T-test was used to compare the LFQ intensities between groups and those with p -values ≤ 0.05 were considered significant. The protein IDs with significant values were subjected to Ingenuity Pathway Analysis (IPA) and the String Database to identify affected canonical pathways and functional protein–protein interaction network. A selection of differentially expressed proteins was ultimately confirmed with immunoblotting as described above.
Male C57BL/6JCr wild-type mice were purchased from Charles River at the age of 10 weeks with a body weight of 25 g. Animals were socially housed with a maximum of eight animals in standard mouse cages (22.5 cm × 16.7 cm × 14 cm) at constant humidity and temperature in a 12/12 h light–dark cycle. Food and water were available ad libitum. Cage enrichment was supplied by a platform, tunnel, and extra cotton sticks. Ex vivo experiments, such as immunohistochemistry (IHC) and co-immunoprecipitation (CoIP), were performed with cerebellar tissue at the age of 10 weeks. All conducted experiments were in compliance with the EU Directive 20,120/63/EU under ECD code 2022–59 after approval by the Animal Ethics Committee of the University of Antwerp.
Immunohistochemistry of frozen murine brain sections (IHC-Fr)
Male C57BL/6JCr wild-type mice were used for immunohistochemistry experiments at the age of 10 weeks. All animals were anesthetized by an intraperitoneal injection of 133.3 mg/kg Dolethal (Vetoquinol; BE-V171692 ), then transcardially perfused for four minutes with 0.1 M phosphate-buffered saline (PBS), subsequently for six minutes with ROTI®Histofix 4% paraformaldehyde solution (pH 7) (Carl Roth; 3105 ) using steady perfusion rate of 12 rpm (2 ml/min). Whole brains were removed from the skull and cut in half along the midline. The two hemispheres were placed in ROTI®Histofix 4% paraformaldehyde (pH 7) (Carl Roth; 3105 ) for two hours at room temperature, washed in PBS (0.01 M; pH 7.4) and transferred in 20% sucrose/PBS for overnight incubation at 4 °C. Tissue samples were embedded in PELCO® Cryo-Embedding Compound (Ted Pella, Inc.; 27300 ) and stored at − 80C. Tissue was cut in sections of approximately 10 µm thickness using the Leica CM1950 Cryostat Microtome (Leica Biosystems, Wetzlar, Germany) and transferred to VWR® SuperFrost® Plus, Adhesion Slides (VWR; 631-0108 ). The sections were washed three times using PBS. After blocking and permeabilization with PBS containing 0.05% thimerosal, 0.01% NaN 3 , 0.1% BSA, 1% Triton X-100 and 10% normal horse serum, sections were incubated overnight with primary ADNP antibody (Abcam; ab300114 ) or SIRT1 (Abcam; ab189494 ) antibody 1:500 diluted in the blocking/permeabilization buffer at room temperature. Tissue sections were washed six times with PBS, followed by a 4-h incubation with Cy3-conjugated Fab Fragment donkey anti-rabbit (Jackson ImmnoResearch Europe Ltd; 711-167-003 ) antibody 1:2000 diluted in PBS containing 0.05% thimerosal, 0.01% NaN3, 0.1% BSA and 10% normal horse serum. After six final washing steps in PBS, nuclei were stained with 5 µg/ml DAPI for 5 min, followed by three washes in PBS. Immunostained cryosections were mounted in Citifluor™ AF1 Mountant Solution (Electron Microscopy Sciences; 17,970–100 ). Confocal images were obtained using a Leica SP8 confocal scanning microscope (Leica-microsystems, Wetzlar, Germany) equipped with a 405-nm diode laser (to detect DAPI) and a white light laser (WLL) used at 555 nm to visualize Cy3. Images were acquired with a 20 × objective (HC PL APO 20x/0.75 IMM CORR CS2). Acquired images were analyzed in FIJI image analysis freeware [ 33 ]. Nuclei were identified as DAPI + regions after automated thresholding of the smoothed DAPI channel (gaussion blur with kernel size 2).
Co-immunoprecipitation (Co-IP) assay
Proteins were extracted from the wild-type mouse cerebellum using N-PER™ Neuronal Protein Extraction Reagent (Thermo Scientific; 87,792 ), supplemented with 1 mg NAP/Davunetide (MedChemExpress; HY-105066 ) to enhance EB1/EB3 binding [ 22 , 23 ], and subjected to Co-IP analysis with the Pierce™ Co-Immunoprecipitation Kit (Thermo Scientific™; 26149 ) according to the manufacturer’s protocol. Briefly, 10 μg of antibodies of interest EB1 (Abcam; ab53358 ) and EB3 (Abcam; ab157217 ) were cross-linked to 50 μl of AminoLink Plus Coupling Resin. An amount of 1 mg of protein lysate was incubated overnight at 4 °C on an end-over-end shaker (VWR; 444-0503 ). Protein elution was performed in three steps: 10 μL, 35 μl, and 50 μl respectively. The immunoprecipitated materials were subsequently investigated by immunoblotting using the following primary antibodies (Additional file 1 : Table S2): rat monoclonal EB1 (Abcam; ab53358 ), rabbit monoclonal EB3 (Abcam; ab157217 ), rabbit monoclonal ADNP antibody (Abcam; ab300114 ) and SIRT1 (Abcam; ab189494 ). In addition, Pierce™ Control Agarose Resin (crosslinked 4% beaded agarose) was used as negative control (IgG). Upon immunoblotting (see above) proteins were visualized using the SuperSignal™ West Femto Maximum Sensitivity Substrate (ThermoScientific™; 34094 ) after labeling with the appropriate secondary antibody (Agilent) in a 1:2000 dilution.
Motif analysis and molecular docking of ADNP and SIRT1 to microtubule-end binding proteins 1 and 3 (EB1/3)
Motif analysis of murine Adnp (UniProt; Q9Z103), Sirt1 (UniProt; Q53Z05), Eb1 (UniProt; Q61166), and Eb3 (UniProt; Q6PER3) was performed using Eukaryotic Linear Motif (ELM) ( http://elm.eu.org ). Three-dimensional models were either generated with AlfaFold ( https://alphafold.ebi.ac.uk ) or obtained from the AlfaFold database (see above) and used to predict protein–protein interactions between Adnp and Sirt1 with both Eb1 and Eb3 using the ClusPro server (ClusPro2.0)141-144 ( https://cluspro.org ), a widely-used protein–protein docking tool. The top 10 resulting motifs were superimposed in UCFS ChimeraX (version 1.6.1.) to present the most probable binding interaction with Adnp and Sirt1.
Screening RNA sequencing data using a mitophagy gene panel
Mitophagy-related gene signature was obtained by clustering analysis of RNA sequencing data from the ADNP brain autopsy and LCLs of ADNP patients and control lines upon gene set enrichment analysis with a customized gene toolbox [ 34 ] in the Omics playground v2.8.22 (Additional file 1 : Table S5). We confirmed the expression of mitophagy- and mitochondrial-related genes (Additional file 1 : Table S4) using RT-PCR as described above.
Autophagy flux assessment
The autophagy flux was determined in ADNP patient and control LCLs by treatment with 160 nM of bafilomycin A1 (Santa Cruz Biotechnology, sc-2021550) for 2 h. Untreated cells and bafilomycin A1-treated cells were collected by centrifugation, and subjected to western blotting as described above. We detected expression of autophagy markers anti-p62/SQSTM1 (Abcam; ab56416 ) with an 1:2000 dilution and anti-LC3 (Abcam; ab192890 ) with an 1:1000 dilution in untreated and treated conditions to assess the autophagy flux. All western blots were controlled by GAPDH incubation (Cell signaling technology; 4317 ). Image Quantification was performed using ImageJ software. Graphical representation was performed in GraphPad Prism version 9.3.1 using a 2-way ANOVA with Šídák's multiple comparisons test.
Live cell imaging: mitochondrial redox state and subcellular localization
Intact fibroblasts of control and ADNP patients (n = 2) were seeded at a density of 4 × 10 6 cells in a 6-well plate for live cell imaging and subsequently stained with 250 nM fluorescent probe MitoTracker® Red CM-H2XRos (Invitrogen; M7513 ) according to the manufacturer’s protocol to assess mitochondrial redox state and subcellular localization. The redox state of mitochondria is determined by the levels of NAD + /NADH, FAD/FADH2, NADP + /NADPH, glutathione/glutathione disulfide (GSH/GSSG) and reactive oxygen species (ROS), which reflect mitochondrial metabolic activity and overall fitness. If the electron transport chain is compromised or there is an imbalance in the redox state, leading to increased ROS production. When CM-H2XRos enters the mitochondria, they are oxidized depending on the relative amount of reactive oxygen species (ROS) present in the mitochondria. The oxidation leads to a change in the fluorescence properties of the red dye. The emitted red fluorescence signal was measured with a multimode microplate reader (Tecan Spark™) and analyzed using the Spark Control™ V3.2 application. The fluorescent signal was statistically quantified using an unpaired Student T-test assuming equal variances in GraphPad Prism 9.3.1. Additionally, fibroblasts were imaged with the Olympus CKX53 fluorescence microscope (Olympus, Antwerp, Belgium) to visualize subcellular localization of the mitochondria with better resolution.
Determination of mitochondrial DNA copy number
Total DNA was isolated from ADNP LCLs and skin fibroblasts as compared to their controls using the DNeasy Blood and Tissue Kit (Qiagen; 69504 ) according to the manufacturer’s instructions. Mitochondrial DNA copy number (mtDNA-CN) was determined using RT-PCR. Briefly,the cycle threshold (Ct) value of a mitochondrial-specific ( tRNAleu ) and nuclear-specific ( B2M ) target were determined in triplicate for each sample using the following primers: tRNA LEU -Fwd: 5′-CACCCAAGAACAGGGTTTGT-3′ and tRNA LEU -Rev: 5′-TGGCCATGGGTATGTTGTTA-3′ and B2M-Fwd: 5′-TGCTGTCTCCATGTTTGATGTATCT-3′ and B2M-Rev: 5′-TCTCTGCTCCCCACCTCTAGGT-3′. The difference in Ct-values (ΔCt) for each replicate represents a raw relative measure of mtDNA-CN.
Seahorse XF cell mito stress test
ADNP patient and unrelated sex- and age-matched control fibroblasts (n = 2) were cultured on Seahorse XFp miniplates with a density of 4 × 10 4 cells (Agilent Technologies; 103725-100 ) and incubated overnight at 37% O 2 /5% CO 2 . Prior to the Seahorse XS Cell Mito Stress Test assay, the fibroblast medium was replaced with Seahorse XF RPMI medium (pH 7.4) (Agilent Technologies; 103576-100 ), supplemented by 1.0 M Seahorse XF Glucose Solution (Agilent Technologies; 103577-100 ), 100 mM Seahorse XF Pyruvate Solution (Agilent Technologies; 103578-100 ), and 200 mM Seahorse XF L-Glutamine Solution (Agilent Technologies; 103579-100 ). The drug ports of the sensor cartridge were loaded with 1 µM Oligomycin (port A), 0.7 µM Carbonyl cyanide-4- (trifluoromethoxy)phenylhydrazone (FCCP) (port B), and 0.5 µM Rotenone/Antimycin A (Rot/AA) (port C). Next, cells seeded in the Seahorse XF HS Miniplates, together with the sensor cartridge, were loaded into the Seahorse XF HS Mini Analyzer (Agilent; S7852A) and subjected to the Agilent Cell Mito Stress Test assay (Agilent; 103010-100) to determine the real-time oxygen consumption rate (OCR) for 1.5 h. First, the baseline respiration was measured (basal OCR) prior to mitochondrial perturbation by sequential injection of 1.5 µM oligomycin (a complex V inhibitor to decrease the electron flow through electron transport chain (ETC)); 3 µM FCCP (the uncoupling agent to promote maximum electron flow through ETC), and a mixture of 0.5 µM Rotenone/Antimycin A (complex I and complex II inhibitors, respectively, to shut down the mitochondria-related respiration). All compounds were included in the Seahorse XFp Cell Mito Stress Test Kit (Agilent; 103010-100 ). The data was analyzed using Agilent Seahorse analytics (Agilent Seahorse Analytic). Statistical analysis was performed in GraphPad Prism 9.3.1 using an unpaired student T-test assuming equal variances.
Clinical presentation
The patient was born prematurely, at 32 weeks of gestational age, from healthy, non-consanguineous parents. His birth weight was 1790 g, the Apgar score was 10/10. An intracranial hemorrhage grade III was diagnosed. Clinical reports showed that the patient presented with motor delays, developmental delays, autism spectrum disorder, hypotonia, and small genitalia. His parents also reported visual impairments, feeding and eating problems, as well as sleep disorders. Phenotypically, the patient presented with a prominent forehead and eyelashes, downward slanting eyes, malformed ears, wide nasal bridge, broad and long philtrum, large mouth with thick lower vermillion, pointed chin and widely spaced teeth (Fig. 1 A, B), all well-defined characteristics described in a cohort of 78 Helsmoortel–Van der Aa patients [ 2 , 4 ] (Additional file 1 : Table S6). At the age of 2.5 years, he developed an upper respiratory tract infection complicated with hepatitis and seizures. He was transferred to ICU where supportive treatment and plasmapheresis were started. Liver biopsy showed extensive necrosis of parenchyma and moderate cholestasis. MRI showed diffuse cortical atrophy of the brain parenchyma, marked reduction in volume of white matter as well as gliosis in both frontal and temporoparietal lobes that could indicate the sequelae of acute hepatic encephalopathy. He developed refractory generalized epilepsy and received a combination treatment of antiepileptic drugs, e.g., carbamazepine, oxcarbamazepine, levetiracetam, clonazepam, clobazam and topiramate. During his lifespan, he underwent two liver transplantations and received immunosuppressants. Following the second liver transplant, at the age of six years and three months old, the child passed away because of multiple organ failure. An autopsy was performed, and various tissue samples were donated with informed consent. Molecular testing had indicated that the patient was negative for any inheritable metabolic disorders. Whole-exome sequencing (WES) of the patient’s blood revealed a heterozygous de novo duplication of adenine at position 1676 in the ADNP gene at position chr20:50,893,037-50,893,039 (RefSeq isoform ENST00000621696.5 Human GRCh38/hg38). The mutation was confirmed by Sanger sequencing (Fig. 1 C). It converts the histidine (His) residue at position 559 to glutamic acid (Gln), leading to a frameshift mutation with a premature stop codon two amino acids downstream (Fig. 1 D, E).

Identification of a heterozygous de novo mutation in the ADNP gene. ( A ) Facial photograph of the six-year-old child ( https://www.adnpfoundation.org/ ). ( B ) Schematic representation of the clinical manifestation of the patient with Helsmoortel–Van der Aa syndrome, including autism, severe ID, and epilepsy. ( C ) DNA sequencing chromatogram of control and patient alleles, confirming a heterozygous nucleotide duplication (c.1676duplA) in the ADNP gene, ( D ) replacing the histidine at residue 559 with glutamic acid with a frameshift of two amino acids and introduction of a stop codon (p.His559Glnfs*3). ( E ) ClustalW alignment across multiple species of ADNP amino acids 520–580. Almost all residues of the ADNP protein are highly conserved amongst vertebrates. The arrow (↓) indicates the species-conserved histidine (H) residue, which is altered in the patient to a glutamic acid (Q) residue. The asterisk (*) indicates positions which have a single, fully conserved residue. A colon (:) indicates conservation between amino acid residues of similar properties
Cerebellar tissue, known for its highest ADNP expression [ 35 ], allowed to validate the presence of ADNP mRNA and protein in autopsy material by performing an expression analysis using real time reverse-transcription PCR (RT-PCR) and Western blotting. To investigate wild-type ADNP mRNA levels, we designed a primer set at the 3’ region of exon 6 (corresponding to the C-terminal portion of the protein). Here, a significant two-fold increase in the total ADNP levels was observed in the patient compared to the control subject ( p = 0.0001; ***), consistent with findings in our RNA sequencing described below (Fig. 2 A). Attempts to quantify the 5’ end of the transcript were not successful, suggesting partial mRNA degradation. At the protein level, we tested endogenous ADNP levels in the human brain using extensively validated C-terminal and N-terminal ADNP antibodies [ 29 ]. We were able to detect wild-type ADNP levels (150 kDa) in the control brain, but not in the patient using both antibodies (Fig. 2 B, C). To investigate the co-expression of the full length and mutant protein, we co-transfected wild-type and p.His559Glnfs*3 mutant N-DYKDDDDK (Flag®) expression vectors in HEK293T cells. Co-expression of wild-type and mutant ADNP demonstrated the presence of the wild-type protein (150 kDa) together with a truncated mutant protein (63 kDa) using an N-terminal antibody, mimicking the expected expression in the patient. C-terminal antibody incubation resulted in the detection of the wild-type ADNP (150 kDa) exclusively. Together, these findings confirm a molecular weight of ADNP (150 kDa), above its calculated molecular weight of 123 kDa, but show instability of the protein in post-mortem brain material of the patient.

ADNP expression analysis of the cerebellum. ( A ) RT-PCR showing a significant increase in ADNP mRNA levels in the patient cerebellum compared to an age-and sex matched control subject (*** p = 0.0001; unpaired student T-test). Gene expression values were normalized with three stable reference genes, i.e., β-Actin ( BACT ), β-2-Microglobulin ( B2M ), and Ubiquitin C ( UBC ). (B) ADNP protein expression analysis using an N-terminal antibody. Western blotting showed the presence of wild-type ADNP (150 kDa) in overexpression lysates with presence of the truncated protein. However, expression was absent in the patient as compared to the control, where wild-type ADNP could be visualized. (C) ADNP protein expression analysis using a C-terminal antibody. Western blotting showed the presence of wild-type ADNP (150 kDa) in overexpression lysates, as well as in the control subject, but not in the patient. GAPDH was used as a loading control for normalization
To study the molecular impact of the patient mutation, we performed in silico modeling of the wild-type ADNP protein (UniProt; Q9H2P0) and p.His559Glnfs*3 mutant using AlfaFold. Here, the structure of the wild-type protein demonstrated the DNA-binding homeobox domain in proximity to the bipartite NLS sequence, whereas the neuroprotective NAP motif resides at the surface of the protein, being partially occluded by flexible intrinsically disordered regions (IDRs) and low-complexity regions (LCRs) located near the C-terminus, suggestive for a role for protein–protein interactions [ 36 , 37 ]. Moreover, the eIF-4E binding motifs and the glutaredoxin active site are centrally positioned in the core of the wild-type protein, assembling several of its zinc finger motifs (Fig. 3 A). The p.His559Glnfs*3 mutant truncates the NLS region, impairing nuclear transport [ 11 ]. Moreover, downstream protein domains, including the DNA-binding homeodomain and the HP1 binding motif are also lost as a result of the truncating mutation. Overall, the p.His559Gln*3 mutant lacks some of the IDRs but has a similar structural confirmation compared to the wild-type protein (Fig. 3 B). Subsequently, we examined stable ADNP protein levels in several subcellular compartments including the cytoplasm, nucleus with chromatin-enriched proteins, and the cytoskeleton in HEK293T overexpression lysates. In the cytoplasm, we detected wild-type (150 kDa) and mutant (63 kDa) ADNP using an N-terminal antibody showing no significant difference in expression levels ( p = 0.71; ns). In the chromatin-bound fraction, we visualized the wild-type and mutant protein with a significant decrease of mutant protein levels ( p = 0.03; *). Moreover, we demonstrated the expression of mutant and wild-type ADNP in the cytoskeletal protein fraction. However, we did not observe a significant difference ( p = 0.42; ns) in the expression of the mutant compared to the wild-type protein (Fig. 3 C).

The ADNP patient mutation impairs expression in the chromatin-enriched protein fraction. ( A ) 3D protein structure representation of the wild-type ADNP glutaredoxin active site (pink), NAP octapeptide sequence (fuchsia), eIF-4E interaction motif (blue), nuclear localization signal (dark cyan), homeobox domain (blue violet), and HP1 interaction motif (purple). The NAP domain (fuchsia) presents at the surface of the protein. ( B ) The nuclear localization signal-truncating p.His559Glnfs*3 mutant shows loss of the HP1-binding motif and DNA homeobox domain. ( C ) N-terminal ADNP detection in different subcellular compartments normalized to their protein fraction-specific loading controls. Detection of wild-type N-DYKDDDDK (Flag®)-tagged ADNP shows a molecular weight of 150 kDa. The p.His559Glnfs*3 mutant showed a lower molecular weight of 63 kDa. Cytoplasmic enrichment shows expression of wild-type ADNP (150 kDa) and the mislocalized p.His559Glnfs*3 mutant (63 kDa) with no difference in expression ( p = 0.71; ns). Chromatin-enriched fraction demonstrated partial loss of mutant ADNP levels compared to wild-type ADNP, showing a dramatic decrease in expression ( p = 0.03; *). Cytoskeletal fraction is enriched for wild-type ADNP and the p.His559Glnfs*3 mutant, with no significant difference in expression ( p = 0.42; ns). GAPDH (cytoplasmic fraction), histone H3 (chromatin-bound fraction), and β-actin (cytoskeletal fraction) were used as loading controls. Statistical analysis of the subcellular fractionation immunoblots was performed using an unpaired two-tailed student T-test, assuming equal variances
Genome-wide methylation analysis of the cerebellum demonstrates abnormalities of the cytoskeleton and autophagy together with an aberrant transcription factor function of ADNP during development
As methylation signatures are robust and even conserved in ancient DNA [ 38 ], we decided to start our exploration by performing an EPIC BeadChip array on the cerebellum of the died ADNP patient and an age-matched control brain. Here, we show enrichment of 6289 CpG probes with a minimum 20% difference in methylation in the ADNP patient compared to the control. Specifically, we identified 2394 CpG probes showing hypermethylation (Δβ > 0.2), whereas a vast amount of 3895 CpG probes were hypomethylated (Δβ < −0.2). In addition, 1547 hypermethylated gene probes could be annotated to 1162 genes, while 2500 hypomethylated gene probes were associated with 1842 genes (Additional file 3 : Data S1), indicating a Class I episignature [ 18 , 19 ], extending findings from peripheral blood to the human brain for the first time (Fig. 4 A). Next, we confirmed a selection on genes prioritized for methylation in the 5’UTR, 3’ UTR and transcription start site (TSS) together with associations to autism or other Helsmoortel–Van der Aa syndrome-related clinical features. We selected the hypermethylated genes OTX2, SLC25A21, and DNAJ6 and the hypomethylated genes COL4A2, MAGI2, and CTNND2 for pyrosequencing. Here, we could confirm a higher percentage of CpG methylation in the patient for OTX2 (56%), SLC25A21 (86%), and DNAJ6 (85%) compared to the control subject. Respectively, we could also demonstrate a lower percentage of CpG methylation in the patient for COL4A2 (1%), MAGI2 (2%), and CTNND2 (3%) (Fig. 4 B). Next, we performed functional annotation of the hyper- and hypomethylated genes using Metascape. Enriched biological processes and GO terms included actin filament-based processes, cell adhesion, nervous system development, muscle contraction, brain development, the WNT signaling pathway, regulation of membrane potential, and synaptic transmission amongst others (Fig. 4 C). Functional enrichment analysis for protein–protein interactions was predicted for ADNP using the STRING database. We identified four suggested interactions of ADNP with WDFY3, UBR5, FAT1, and NFIA, which play a role in autophagy of the mitochondria, protein ubiquitination, macro-autophagy, autophagosome and autolysosome formation (Fig. 4 D). Given the role of Adnp as a putative transcription factor [ 35 , 39 ], we performed a transcription factor enrichment of both hyper- and hypomethylated genes. Here, we identified a module of 44 co-expressed genes, which were subsequently inserted in CytoScape using the IRegulon function for TF enrichment (Additional file 4 : Data S2). We observe a stronger enrichment of TFs associated with hypomethylated genes (red) than hypermethylated genes (blue) and shared TFs (green). Among the upregulated TFs associated with hypomethylated genes presented pluripotency and cell fate-determining genes such as POU2F1 , TEAD2 , SOX1/4 , GATA1/2/3/5/6, PAX4/6, NANOG, and NEUROD1 , as well as chromatin modifiers like YY1, SIN3A and ADNP itself. On the other hand, the downregulated TF cluster associated with hypermethylated genes was also enriched for PAX and SOX -related genes, indicating abnormal lineage specification of neuronal progenitor cells. The shared TF cluster showed presence of HNF1A , a gene controlling expression of several liver-specific genes (Fig. 4 E). Our genome-wide cerebellar methylation analysis indicates strong molecular evidence for a deregulated function of ADNP as a transcription factor, impacting lineage specification and genes implicated in brain development.

ADNP methylation signature in the juvenile post-mortem cerebellum. ( A ) Genomic scatter plot indicating the hypermethylated genes (Δβ > 0.2) of the patient (red), the hypomethylated genes (Δβ < −0.2) of the patient (blue). The chromosomal positions of the genes are shown on the x-axis. ( B ) Pyrosequencing confirmation of a subset of hyper- and hypomethylated genes. Hypermethylated genes, e.g., OTX2 , SLC25A21 and DNAJ6 , show increased CpG methylation in the patient, whereas hypomethylated genes, e.g., COL4A2 , MAGI2 and CTNND2 , present with a nearly absent percentage of CpG methylation. ( C ) Metascape functional annotation of biological processes. Hyper- and hypomethylated genes cluster in associated processes such as the actin cytoskeleton and nervous system developmental disorder amongst others. ( D ) Predictive String v11.5 protein–protein interaction analysis of ADNP. The proteins are indicated as nodes with interconnecting lines representing the interaction. ADNP is surrounded by protein regulating specific autophagy-related processes and protein ubiquitination. ( E ) Transcription factors (TFs) enriched in patient cerebellum of hyper- and hypomethylated gene co-expression. TFs associated with hypermethylated genes are represented in blue, while the TFs associated with the hypomethylated genes are depicted in red. TFs shared amongst the overlapping genes are shown in green. ADNP was identified as the top transcription factor controlling the hypomethylated genes (black box)
RNA sequencing substantiates downregulation of the WNT signaling pathway and autophagy defects in cerebellar autopsy tissue
To determine differential expression beyond methylation differences, we performed bulk transcriptome sequencing of cerebellar tissue of the ADNP autopsy. As RNA is much less stable over time, we first performed an extensive quality control by evaluating total RNA purity and integrity (see experimental methods). Using bulk mRNA sequencing, we determined the gene ratio (patient/control) using the NOISeq algorithm, a non-parametric method for comparing samples without biological replicates, reporting the log2-ratio of the two conditions (M) and the value of the difference between the conditions (D) [ 40 ]. We tested for differential expression across all 7659 genes that appeared in our data set (Additional file 5 : Data S3). In line with the observation of an excess of hypomethylated CpG probes, we observed an excess of upregulated genes. Using a significance cut-off equivalent to, p value < 0.05, FDR = < 0.05, and a biologically meaningful (M-value) log2FC > 0.5, we found 514 downregulated and 1520 upregulated genes with differential expression (Fig. 5 A). Gene expression alterations in the ADNP cerebellum were notable with the majority of genes presenting with an M-value < 5. Gene ontology (GO) enrichment revealed downregulation of glutamatergic synaptic transmission, abnormal cardiac muscle cell conductivity, and nervous system development, whereas cytoskeleton dynamics were upregulated. A remarked enrichment of immune system-related responses was observed that are potentially related to the patient’s immunosuppressant treatment (Fig. 5 B). We confirmed a selected set of genes with RT-PCR, including the RNA-methylation gene METTL3 ( p = 0.005; **), autophagy inducer BECN1 ( p < 0.0001; ****), and WNT signaling ligand CTNNB1 ( p = 0.001; **) (Fig. 5 C). To better interpret the differential expression in the ADNP brain, we compared the transcriptome analysis of the autopsy with the differential expression observed in immortalized LCLs of multiple patients with different ADNP mutations. We tested for differential expression across approximately 10,000 protein-coding transcripts that appeared in our data set (Additional file 6 : Data S4). Using the exact cut-off criteria as in the autopsy, we found 1730 downregulated and 3278 upregulated genes with differential expression, indicating that the ADNP mutations rather induce gene upregulation (Fig. 5 D). Fast Gene Set Enrichment Analysis (fgsea) identified similar molecular pathways as identified in the autopsy (Fig. 5 E). We confirmed a subset of five genes with RT-PCR, including the heterochromatin marker and ADNP-interacting gene CBX3 ( p = 0.01; *), WNT signaling member WNT10A ( p = 0.003; **), actin-cadherin mediator CTNNAL1 ( p = 0.003; **) as well as nonsense mediated decay members SMG5 (p = 0.0002; ***) and UPF3B ( p = 0.005; **) (Fig. 5 F).

Cerebellar and lymphoid gene expression changes are associated with different ADNP mutations. ( A ) Volcano plot of differentially expressed genes (DEGs) in the ADNP cerebellum using the NOISeq algorithm, representing the effect size M (log2 ratio) and D (difference between conditions) values. The DEG are shown in blue. ( B ) Gene set enrichment analysis of all DEGs in gene ontology (GO), biological processes (BP) and molecular function (MF) reveals specific Helsmoortel–Van der Aa syndrome-related pathways in the ADNP brain. ( C ) RT-PCR showing a significant reduction of METTL3 , BECN1 and CTNNB1 mRNA levels in the ADNP cerebellum compared to the age-matched control subject. Expression values were normalized using the housekeeping genes ACTB , B2M and UBC . Data was analyzed using an unpaired student T-test. ( D ) Volcano plot of DEGs in the patient LCLs using the DESeq2 package, displaying the significance (-log10q) and effect size (log2FC). The DEG are shown in blue. ( E ) Functional gene set enrichment of GO and BP using differentially expressed genes in the ADNP LCLs as compared to age- and sex-matched controls. UMAP clustering of gene sets colored by standard deviation, variance, or mean fold-change in patient LCLs shows clear downregulation of the WNT, Hedgehog and Notch signaling pathways (marked in a red box), impairing proper neuronal development. Downregulated genes, blue; upregulated genes, red. ( F ) RT-PCR showing a significant increase of CBX3 , CTNNAL1, SMG5 and UPF3B together with a significant decrease in WTN10A mRNA levels in patients versus control LCLs. Expression values were normalized using the housekeeping genes GAPDH , RPL13A and SDHA . Data was subsequently analyzed with a Mann–Whitney U test for unpaired measures
To investigate the potential impact of the ADNP mutation in the human brain, we intersected the DEGs identified in both data sets (Additional file 7 : Data S5), which revealed an overlap of 241 genes between the ADNP autopsy brain and LCLs (Fig. 6 A). We observed a striking resemblance for biological relevance of genes involved in endoderm specification IGFBP2 (brain, * p = 0.03; LCL, * p = 0.04), canonical WNT signaling WNT2 (brain, * p = 0.01; LCL, ** p = 0.01), mitochondrial transporter SLC25A25 (brain, * p = 0.02; LCL, * p = 0.03), autophagy regulation RUBCN (brain, **** p < 0.0001; LCL, * p = 0.003), hematopoietic stem cell differentiation RUNX1 (brain, ** p = 0.001; LCL, *** p = 0.001), N 6 -adenosine-methylation METTL3 (brain, ** p = 0.005; LCL, **** p < 0.0001), and bone and teeth development BMP6 (brain, ** p = 0.002; LCL, * p = 0.04) (Fig. 6 B, C). In conclusion, these robust gene expression changes related to the nervous system and morphogenesis underline a regulating role of ADNP in the human brain and blood of patients, confirmed by salient pathways including the WNT signaling, autophagy, and bone development together with involvement in processes such as hematopoietic stem cell differentiation and unexpected RNA methylation.

Meta-analysis of the transcriptomic signature identified in the ADNP brain and lymphoblasts. ( A ) Venn diagram representing the exact amount of DEGs in the human ADNP cerebellum and lymphoblastoid data sets, converging to an amount of 241 overlapping genes. ( B ) RT-PCR showing a significant decrease of IGFBP2 , WNT2, SLC25A25 together with a significant increase in RUBCN and RUNX1 mRNA levels in patient brain and patient LCLs as compared to their age-matched controls. Note a difference in mRNA expression of BMP6 and METTL3 amongst brain tissue and LCLs. Expression values were normalized using the tissue-specific housekeeping genes (mentioned above). Data was subsequently analyzed with an unpaired student T-test (brain) or Mann–Whitney U test for unpaired measures (LCLs). ( C ) Correlation of DEGs from the NOIseq and DESeq2 analysis (RNA-seq) and RT-PCR confirmations with their functional cellular role. Selected genes are represented with their Log2FC and compared for overlap with RT-PCR. Arrow upwards, upregulation; arrow downwards, downregulation
Shotgun proteomics links chromatin remodeling to autophagy in the ADNP autopsy brain
As post-transcriptional regulation can further increase variation in gene expression levels [ 41 ], proteome analysis was performed by label-free quantitation (LFQ) mass spectrometry on the cerebellum to study the effect of the c.1676duplA/p.His559Glnfs*3 ADNP mutation at protein expression level. Chromatographic conditions between different runs were highly reproducible, resulting in a strong correlation between LFQ intensities and technical replicates (Additional file 8 : Data S6). Overall, we detected approximately 1522 protein groups per sample under a 1% false-discovery rate (FDR) with fixed modifications of carbamidomethylation (C), deamidation (QN) and oxidation (M). Moreover, we identified 4552 proteins with more than two unique peptides, respectively 988 proteins with at least two unique peptides, and 1477 with one unique peptide. Next, we used MetaboAnalyst 5.0 to quantify differences detected in patient versus control cerebellum. Among the 2455 quality-filtered proteins, we detected 492 proteins with a differential expression (Additional file 9 : Data S7), of which 224 proteins were significantly downregulated, while 268 proteins showed a significant upregulation in the post-mortem patient cerebellum (two-tailed student T-test; padj < 0.05). Next, we plotted the top 10 downregulated (represented in blue) and upregulated (represented in red) proteins identified in patient versus control brain, showing a clear upregulation of the major ADNP-interacting protein heterochromatin Protein 1 homolog beta (CBX1/HP1β), amongst others, indicating that ADNP is able to somehow affect the expression of its direct interaction partner (Fig. 7 A). Subsequently, we applied immunoblot experiments to confirm the downregulation of β-catenin and BECN1 protein levels in the patient brain, in line with its decreased transcription levels. Surprisingly, we also observed differential expression of an additional autophagy marker, MAP1LC3A, in the ADNP brain consistent with aberrant autophagy defects in our transcriptome data (Fig. 7 B). Clustering of the differentially expressed proteins (DEPs) in canonical pathways using IPA indicated a decreased activity of mitochondrial oxidative phosphorylation, sirtuin signaling and RhoA signaling. In contrast, IPA predicted an increase in EIF2 signaling, spliceosomal cycle and protein kinase A signaling in the patient. We also observed an enrichment of pathways with no difference in activity, including granzyme A signaling and mTOR signaling, T-helper signaling, and apoptosis (Fig. 7 C). Next, we mapped all DEPs in a functional enrichment analysis and predicted possible protein–protein interactions of ADNP with the identified DEPs as well as with other biologically correlated proteins. Of particular interest, the histone deacetylase sirtuin 1 (SIRT1) in the center of the protein network was found to link various chromatin modifier proteins such as MECP2, ADNP, SMARCC2, HDAC2 including YY1, and chromatin-associating proteins such as CBX1/3, histones H1F0 and H1.2 to autophagy regulators like MAP1LC3A and LAMP1 (Fig. 7 D). In this section, we showed that the proteomic landscape of ADNP brain autopsy material corroborates our transcriptome findings, e.g., upregulation of ADNP-interactor CBX1/HP1β together with a downregulation of β-catenin and BECN1, supported by abnormalities attributed to the WNT signaling pathway and autophagy.

Discovery of SIRT1 interactions in the autistic brain, linking chromatin remodelers to autophagy. ( A ) Volcano plot of differentially expressed proteins (DEPs) in the ADNP patient cerebellum, represented by the significance (−log10( p )) and effect size (log2FC). Proteins with a significant downregulation are shown in blue and those with an upregulation in red. Note the marked difference in expression of the ADNP-interacting protein CBX1/HP1β. ( B ) Immunoblot confirmations of DEPs exposed to their specific antibodies. ( C ) Top 15-ranked canonical IPA pathways of the ADNP brain, represented as z-scored expression values and significance (−log p -value). Activated pathways are presented in orange, respectively lowered activity in blue; no activity pattern available (grey); pathways with no difference in activity (white). ( D ) A predictive protein–protein interaction network was generated by String Version 11.5, integrating associations of a co-expression hub identified amongst the DEPs. Proteins are represented by colored network nodes in relation to each other with SIRT1 fulfilling a centralizing role. The edges illustrate functional associations and the lines between the nodes represent the existence of evidence for associations
ADNP and SIRT1 co-immunoprecipitate with the microtubule end-binding proteins 1 and 3 (EB1/EB3)
Recently, various studies identified an association between mitochondrial dysfunction, autophagy regulation, and autism spectrum disorders [ 42 , 43 , 44 ]. Similarly, our proteomic protein–protein interaction study mapped SIRT1 at the crossroads of chromatin remodelers and autophagic regulators in the ADNP autopsy brain. Besides, SIRT has been discovered to maintain genomic stability [ 45 ], to enhance synaptic plasticity [ 46 ], to suppress inflammation [ 47 ], to fulfill a neuroprotective function [ 48 ], and to positively regulate autophagy and mitochondrial function [ 49 ]. In addition, SIRT1 is known to modulate chromatin structure by activating BRG1, which is a chromatin remodeling interaction partner of ADNP in the SWI/SNF complex [ 9 , 50 ]. Hence, we reasoned that ADNP and SIRT1 may share common regulatory partners in chromatin remodeling and microtube dynamics that regulate autophagy. To further validate a direct protein interaction of ADNP and SIRT1 in the human brain, co-immunoprecipitation (co-IP) experiments were performed. However, due to the instability of the ADNP protein, co-IP experiments were not successful. Therefore, we alternatively demonstrated the subcellular localization of Adnp and Sirt1 by immunostaining the cerebellum dissected from male C57BL/6JCr wild-type mice as a model for the human condition. Here, cerebellar cryosections were immunostained with primary monoclonal ADNP and SIRT1 antibodies (Cy3 red fluorescent signal), and nuclei were counterstained with DAPI (blue). Adnp expression was predominantly detected in the nucleus, with occasional weak cytoplasmic signals, visualized by the overlap of the red Adnp signal and blue DAPI counterstaining. In contrast, Sirt1 was predominantly situated in the cytoplasm of Purkinje cells in the cerebellum, with occasional nuclear immunoreactivity (Fig. 8 A). Indeed, an indirect interaction of ADNP and SIRT1 was shown in SH-SY5Y cells which could not be validated in human induced pluripotent stem cells (hIPSC)-derived neuronal differentiated cells [ 16 ]. Therefore, we next investigated the potential indirect interaction of Adnp and Sirt1 through the EB1/EB3 proteins in murine cerebellar brain lysates with a co-immunoprecipitation assay. During this process, we performed stringent washing steps using high detergent buffers to prevent false-positive binding. In addition, we also controlled each western blot with GAPDH, whose intensity was absent after co-immunoprecipitation of the bait protein. We observed specific co-immunoprecipitation of Adnp (150 kDa) and Sirt1 (100 kDa) in the presence of both EB1 (30 kDa) and EB3 (32 kDa) antibodies. IgG non-reactive beads were used as a negative control, showing no immunoreactivity of Adnp, Sirt1 together with EB1 and EB3 in the eluted fraction (Fig. 8 B). To better understand the physical connections between Adnp (UniProt; Q9Z103) and Sirt1 (UniProt; Q53Z05), we applied a eukaryotic linear motif (ELM) analysis to unravel shared motifs (Fig. 8 C). Interestingly, as partially shown before, we identified a series of common interaction motifs, including (1) SxIP motif for Adnp (aa 354–360, NAPVSIP, p = 0.01) and similar SSIP for Sirt1 (aa 440–448, VALIPSSIP, p = 0.0002), (2) SH3-domains for Adnp (i.e., aa189–195, FQHVAAP, p = 0.01) and Sirt1 (i.e., aa 506–511, PPRPQK, p = 0.001), and (3) 14–3–3 motifs for Adnp (i.e., aa 16–20, RKTVK, p = 0.004) and Sirt1 (i.e., aa 333–342, RNYTQNIDTL, p = 0.004). The presence of a SxIP motif is a unique feature ascribed to both ADNP and SIRT1, as only 42 protein have been identified by mass-spectrometry based methods to contain this conserved motif [ 51 , 52 ]. Original studies have identified the microtubule-end binding proteins EB1 and EB3 as interaction partners of ADNP through its SxIP motif [ 16 , 23 , 27 ]. Along the same line, we predicted a physical interaction between Adnp, Sirt1 and EB1/EB3 in silico via 3D-molecular docking. Upon ranking the models according to the amount of interacting amino acids, the top 10 docking interactions were derived from ClusPro and processed in ChimeraX. Molecular docking revealed possible Adnp (blue) binding to both microtubule-end binding proteins EB1 (left, violet) and EB3 (right, pink) via amino acids 358–360, corresponding to its SxIP motif. In addition, Sirt1 was predicted to interact with EB1 (left, violet) and EB3 (right, pink) proteins through its similar SSIP motif at amino acid position 446–448 (Fig. 8 D). In conclusion, our findings suggest that Adnp and Sirt1 might indirectly co-immunoprecipitate in the presence of the EB1/EB3 proteins via the SxIP motif for ADNP, respectively SSIP for SIRT1.

ADNP indirectly interacts with the histone deacetylase enzyme SIRT1 through the microtubule end-binding proteins EB1/EB3. ( A ) Adnp and Sirt1 immunostaining (red Cy3 fluorescence) in cryosections of the murine cerebellum was assessed by confocal scanning microscopy. Adnp was mainly observed in the nucleus and Sirt1 was mostly located in the cytoplasm. ( B ) Co-IP assay of Adnp and Sirt1 in the murine cerebellum. IP-competent EB1 and EB3 antibodies were crosslinked to agarose beads and sequentially eluted in fractions (input, In; flow-through, Ft; three consecutive washes, w1-w3; and elution, E). All fractions were analyzed by western blotting for Adnp, Sirt1, EB1, and EB3. IgG non-reactive beads were used as a negative control. GAPDH has been used as loading control for all western blots, and critical assessment of the accuracy of the Co-IP method. ( C ) ELM analysis identified shared motif sequences of Adnp and Sirt, including 14–3–3 motifs (green), SxIP motif (blue), SH3 domains (orange), and the SSIP motif (violet). ( D ) In silico 3D-molecular docking of Adnp (SxIP motif) to EB1/3, left (purple) and right (pink) respectively to Sirt1 in the panel below (SSIP motif) to EB1/3, left (purple) and right (pink)
The ADNP-EB1/EB3-SIRT1 complex regulates mitochondrial autophagy (mitophagy) and respiratory functions
RNA sequencing data of the ADNP brain autopsy was subsequently analyzed for the enrichment of specialized pathways associated with autophagy [ 53 , 54 ], neuroprotection [ 55 , 56 ], and mitochondrial biogenesis [ 57 ] with a costomized gene toolbox [ 34 ]. By this approach, we found enrichment of genes involved in mitophagy (Additional file 1 : Table S5), an autophagy-dependent process regulating mitochondrial homeostasis [ 42 ]. Similarly, RNA sequencing data of the ADNP patient LCLs also revealed enrichment of mitophagy-related genes e.g., ubiquitin Specific Peptidase 15 ( USP15 ), mitochondrial distribution and morphology regulator 1 ( MSTO1 ), mitochondrial fission regulator 2 ( MTFR2 ), apoptosis inducing factor mitochondria associated 1 ( AIFM1 ), Mitofusin 2 ( MFN2 ), Beclin 1 ( BECN1 ), and mitochondrial Elongation Factor 1 ( MIEF1 ) (Fig. 9 A). RNAseq results of nine DEGs were further validated by RT-PCR, including MFN2 ( p < 0.0001; ****), MAPK1 ( p = 0.01; *), BECN1 ( p < 0.006; **), MCL1 ( p = 0.001; **), USP15 ( p = 0.0002; ***), USP8 ( p = 0.02; *), TBK1 ( p = 0.03; *), UBE2N ( p = 0.004; **), and MTFR2 ( p = 0.002; **) (Fig. 9 B). Since we obtained two reciprocally regulated mitophagy gene clusters in the human brain (Additional file 10 : Data S8), we next determined the autophagic flux [ 58 , 59 ] by Bafilomycin A1 treatment in LCLs derived from ADNP patients and sex-age-matched control subjects followed by western blot detection of p62 and LC3 (Additional file 11 : Data S9). Prior to Bafilomycin A1 treatment, we detected a non-significant increase in levels of p62/SQSTM1 ( p = 0.38; ns) and LC3 expression ( p > 0.99; ns) in patient-derived LCLs. Following Bafilomycin A1 treatment, the expression of p62 ( p = 0.18; ns) increased in the ADNP deficient cell lines, although non-significant. However, the levels of LC3 ( p < 0.0001; ****) showed a remarkable increase in the ADNP-patient cell lines, confirming an increased autophagic flux. Given the mitophagy gene signature in LCLs from ADNP patients together with downregulated mitochondrial protein functions identified by LFQ-MS in the human ADNP autopsy brain, we subsequently investigated relative changes in mitochondrial activity and subcellular localization in patient-derived ADNP fibroblasts compared to two unaffected controls using the MitoTracker® Red CM-H2XRos probes. Here, we observed a rather faint fluorescence intensity in the ADNP patient fibroblasts compared to the controls. Furthermore, we did not observed a difference in the subcellular localization of the mitochondria in patient fibroblasts as compared to the control subjects (Fig. 9 C). Relative MitoTracker® Red CM-H2XRos probe fluorescence per cell, indicative of mitochondrial redox activity, was quantified in patient and control fibroblasts using the Tecan Spark™ and normalized to the brightfield cell count. The mitochondria of patient-derived skin fibroblasts showed a remarkable decrease ( p = 0.01; *) in fluorescent intensity compared to the control cells, indicating aberrant mitochondrial activity (Fig. 9 D). To rule out the possible decrease in the mitochondrial copy number in ADNP LCLs and skin fibroblasts, we determined the mtDNA/nDNA ratio (tRNAleu/B2M) by RT-PCR. Since we could not detect a significant difference in the number of mitochondria in ADNP cell lines compared to controls (Additional file 12 : Data S10), we next investigated mitochondrial respiration using the Seahorse analyzer and further addressed the observed mitochondrial dysfunction in patient-derived (blue) and control (red) fibroblasts using the Cell Mito Stress Test to measure changes in oxygen consumption rate (OCR) before/after administration of specific compounds that sequentially affect the different complexes of the mitochondrial respiratory chain. The changes in OCR allowed quantification of several aspects of the mitochondrial respiration. Measurements of the basal respiration showed a significant decrease ( p = 0.04; *) in the patient fibroblasts compared to the control lines, confirming a reduced activity measured in the fluorescent mitochondrial staining. Besides, we also observed decreased values of proton leak ( p = 0.21; ns), ATP-linked respiration ( p = 0.06; ns) and maximal respiratory capacity ( p = 0.28; ns) in the ADNP cell lines, although the difference was not significant. Lastly, we observed no difference in the spare respiratory capacity ( p = 0.84; ns) and non-mitochondrial respiration ( p = 0.79; ns) in patient or control lines (Fig. 9 E, F).

Mitophagy gene signature and mitochondrial impairments in ADNP patient-derived cell lines. ( A ) Expression levels of mitophagy-related genes in ADNP patient LCLs compared to age and sex-matched controls. mRNA sequencing demonstrated an upregulated mitophagy gene signature in patient LCLs as compared to controls. ( B ) RT-PCR showing a significant increase of mitophagy-related genes in ADNP patients versus control LCLs. Expression values were normalized using the housekeeping genes GAPDH, RPL13A and SDHA . Data was subsequently analyzed with an unpaired student T-test assuming unequal variances with a Welch’s multiple testing correction. ( C ) Subcellular mitochondrial distribution using the MitoTracker® Red CM-H 2 XRos fluorescent staining of control subjects (A-B) and ADNP patient (C-D) fibroblasts. The white arrow indicated the unstained nucleus of the fibroblast. ( D ) MitoTracker® red fluorescent signal (RFU) was normalized to brightfield cell count and quantified in patient and control fibroblasts using a multimode microplate reader (Tecan Spark™) at an excitation of 485 nm. A significant decrease ( p = 0.01, student T-test) in red fluorescent signal, corresponding to the mitochondrial activity, was observed in patient-derived fibroblasts (blue) compared to the control cells (red). ( E ) The Seahorse Cell Mito Stress Assay was used to measure changes in oxygen consumption rate (OCR) in patient-derived (blue) and control (red) fibroblasts after different triggers that inhibit or activate mitochondrial respiration. Oligo = oligomycin, a complex V inhibitor to decrease the electron flow through the electron transport chain (ETC); FCCP = Carbonyl cyanide- p -trifluoromethoxyphenylhydrazone, the uncoupling agent to promote maximum electro flow through of the ETC; Rot/AA = rotenone and antimycin, complex I and complex II inhibitors respectively, to shut down the mitochondrial-related respiration. ( F ) Based on the changes in OCR, several aspects of the mitochondrial respiration could be quantified using the Agilent Seahorse analytics software. Statistical significance was calculated using an unpaired student T-test assuming equal variances. Data is shown as mean ± sd. The basal respiration ( p = 0.04, *), proton leak ( p = 0.21; ns), ATP-linked respiration ( p = 0.06, ns), maximal respiration ( p = 0.28, ns), spare capacity ( p = 0.84, ns), and non-mitochondrial respiration ( p = 0.79; ns) were indicated for patients (blue) compared controls (red)
In this study, we investigated the cerebellum of a unique six-year-old male child heterozygous for the ADNP de novo mutation c.1676dupA/p.His559Glnfs*3 who died of multiple organ failure after a second liver transplant to unravel functional biochemical consequences of the mutation in the human brain.
ADNP gene mutations typically result in a syndromic form of autism, co-morbid with ID, termed Helsmoortel–Van der Aa syndrome [ 2 ]. The unique autopsy case analyzed in this report presented with moderate ID, whereas developmental milestones such as sitting up and walking independently, language and bladder training were delayed as reported for almost all children of the Helsmoortel–Van der Aa syndrome. With 83.3% of all patients presenting with gastrointestinal problems, our autopsy case presented with parent-reported feeding and eating problems. He also presented with severe autism typified by avoidant behaviors towards other children as reported in almost 93% of patients. To a lesser extent, the child showed generalized symptomatic epilepsy, which was only reported at low frequency in the population (16.2%). Phenotypically, he presented with key features of the syndrome indicated by a prominent forehead, wide nasal bridge, large mouth, widely spaced teeth, and malformed ears as observed for more than the majority of patients. Symptoms were also correlated to urogenital problems (28%), sleep problems (65.2%), and early teething (71.1%) [ 4 , 60 ]. Clinically, our autopsy patient is thus a bona fide representative of the Helsmoortel–Van der Aa syndrome patient group.
While we have been unable to detect (mutant) ADNP by Western blotting in the autopsy, we do show here a human cerebellar DNA methylation pattern consistent with methylation patterns found in the blood of patients and a transcriptomic profile that bears a significant overlap with the transcriptomic profile of patient cell lines. Proteome analysis of the post-mortem cerebellum also pointed to pathways that have been implicated in the Helsmoortel–Van der Aa syndrome and lead to the identification of ADNP regulatory functions of mitochondria in the human brain. We thus conclude that the ADNP post-mortem cerebellum is valid to study the disorder and further enabled us to confirm earlier observations made in cellular systems and in disease-relevant brain tissue to unravel novel pathways.
The ADNP brain-specific episignature
ADNP-specific defects in chromatin remodeling translate into genome-wide changes in DNA methylation, including differential methylation of various genes involved in cytoskeletal functions, synaptic transmission, nervous system development [ 2 , 8 ], calcium-binding [ 61 , 62 ], and WNT signaling [ 14 ], in part resembling a Class I type episignature of Helsmoortel–Van der Aa syndrome [ 18 , 19 ]. Brain-specific MAGI2 and CTNND2 hypomethylation, similar to hypomethylation detected in peripheral blood of ADNP patients, was confirmed by targeted bisulfite pyrosequencing and illustrates partially conserved ADNP-specific episignatures across brain-blood cell types. Interestingly, CTNND2 dysregulation has been reported in several cases of autism and intellectually disabled patients presenting with behavioral problems and dysmorphic features [ 63 , 64 ], involving changes in autophagy signaling and arborization of the developing dendrites [ 65 ]. Interestingly, our transcription factor (TF) motif enrichment analysis further identified ADNP as the main transcription factor controlling the hypomethylated gene cluster, confirming its central regulatory function during brain development [ 2 , 3 , 8 ].
The ADNP mutation affects lineage specification
ADNP regulates genes participating in embryonic development such as Pax6, Olig2, Sox1, Nestin, and Otx2 [ 8 , 39 ]. Our TF analysis showed enrichment of these lineage specifying genes in the autopsy brain. Pax6 is important for neuronal development [ 8 , 66 ], especially the eye, which could potentially be related to the visual problems observed in our patient cohort (73.6%) [ 4 ]. Olig2 is involved in cell fate of ventral neuroectodermal progenitors and differential expression impaired interneuron differentiation, causing cognitive impairments [ 67 ]. Sox1 and its isoforms regulate embryonic development, male sex determination, and cell fate decisions by acting as an inhibitor of neural differentiation [ 68 ]. Nestin encodes a cytoskeletal protein that is expressed in nerve cells with disturbances causing deficits in motor coordination [ 69 ], a hallmark of the Helsmoortel–Van der Aa syndrome [ 4 ]. The transcription factor Otx2 is involved in the differentiation and proliferation of neuronal progenitor cells and as such affecting brain development, craniofacial and sensory organs, and synaptic plasticity [ 70 , 71 ], which all have been reported to be dysregulated in an Adnp heterozygous knockout and CRISPR/Cas9 mouse models [ 26 , 55 ]. Interestingly, OTX2 hypermethylation was also confirmed by bisulfite pyrosequencing in the ADNP autopsy brain, correlating with abnormal synaptic plasticity, which has previously been demonstrated by immunohistochemical stainings of PSD95 and NMDAR1 in the hippocampal hillus and dentate gyrus of this patient [ 72 ]. Taken together, a mutation in the ADNP gene affects brain methylation and expression of genes involved in brain development, neuronal plasticity, and lineage specification.
The ADNP mutation affects WNT signaling
Consistent with the ADNP-specific episignature pathway enrichment outcome, brain transcriptome changes affected similar pathways, involving downregulation of the WNT signaling pathway [ 14 ], glutamatergic synaptic transmission [ 73 ], cardiac muscle functioning [ 9 ] and nervous system development [ 8 ]. More particularly, we observed a decrease of β-catenin levels, the major transcriptional driver of the WNT signaling pathway, resulting in the downregulation of neuroectoderm developmental genes and defective neurogenesis [ 14 ]. We also found molecular indications for the downregulation of WNT signaling member 10A (WNT10A), a gene essential for tooth morphogenesis [ 74 ]. Interestingly, 71% of children present with premature primary tooth eruptions in the Helsmoortel–Van der Aa syndrome [ 4 , 60 ]. These results were confirmed using bulk mRNA sequencing results of LCLs obtained from several ADNP children showing decreased WNT signaling [ 17 , 60 , 72 ], together with pathways such as Notch [ 14 ] and Hedgehog signaling [ 17 ] affecting embryogenesis and morphogenesis. Another downregulated gene is the RNA-methylating enzyme METTL3 , previously reported to be regulated by the β-catenin/WNT signaling pathway in an autism mouse model [ 75 ], as well as by the FMR1 gene, causative of the autistic Fragile X syndrome [ 76 ].
ADNP plays a suggested role in autophagy and aging
We also observed abnormalities in the autophagy pathway, affecting brain homeostasis [ 77 , 78 ], via downregulation of the autophagy inducer BECN1 in both brain patient brain tissue and brain samples of Adnp deficient mice [ 24 ] as well as in post-mortem schizophrenia brains [ 21 ]. Furthermore, we also demonstrated elevated LC3 levels in the autopsy brain and LCLs after Bafilomycin A treatment. The reduction in BECN1 levels with the spontaneous increase in LC3 protein levels might reflect a compensatory mechanism [ 79 ]. In addition, ADNP was also shown to bind LC3 directly in a human neuroblastoma cell line, and its association was increased in the presence of the NAP (Davunetide) octapeptide [ 78 ]. Furthermore, we observed expression changes in members of the nonsense-mediated decay (NMD) pathway, which trigger the autophagy process [ 80 ]. In fact, changes in mRNA levels of the NMD-members SMG5 and UPF3B have been associated with intellectual syndromic and non-syndromic intellectual disability, autism, childhood onset schizophrenia and ADHD [ 81 , 82 , 83 ]. Interestingly, the Slc12a2 and Slc9a3 family members were shown to be regulated in an age-dependent manner in the hippocampus, cortex, and spleen of Adnp haploinsufficient mice [ 25 ], and early-onset hippocampal tauopathy, a marker for aging and neurodegeneration, was reported in this young subject by an independent study [ 72 ]. RUBCN is a negative regulator of autophagy which also correlates with aging [ 84 ].
HP1beta involvement
Intriguingly, we also detected a significant increase in protein levels of the repressive chromatin ADNP-interactor HP1β in our proteomics experiment exclusively, which is consistent with as yet unpublished observations in the brain of our novel Adnp frameshift mouse model [ 7 ], again highlighting the resemblance between murine and human data and the effects that mutations in ADNP have on the expression of its direct interaction partners [ 9 ].
Novel ADNP interactions in the brain: immune signature, the cytoskeleton and mitochondrial involvement
In our study, pathways involving cytoskeleton dynamics [ 85 ], T-helper cell differentiation and immune-associated pathways [ 86 ] were found to be transcriptionally upregulated. However, it cannot be excluded that the observed immune system malfunctions could be related to the reported multiple organ failure in the patient. Moreover, a substantial part of Helsmoortel–Van der Aa patients presents with (pharmacologically treated) thyroid hormone problems or epilepsy, which may also affect inflammation-related processes [ 4 ]. Upon further downstream protein expression profiling of the ADNP patient versus healthy cerebellum proteome by mass spectrometry analysis, differentially expressed proteins showed enrichment for mitochondrial dysfunction. Mitochondrial dysfunction has been linked to the onset of autism [ 87 , 88 ] and epilepsy [ 89 , 90 ] by multiple studies, both key features present in the clinical presentation of the current case. Besides, we also observed significant downregulation of cytoskeletal functions and sirtuin signaling. Of special note, ADNP associates with cytoskeletal microtubules through its SxIP motif and the microtubule end binding proteins 1 and 3 (EB1/EB3) in differentiated neurons [ 22 ] with microtubule deficits of human ADNP mutants in cell cultures (e.g., p.Ser404*, p.Tyr719*, and p.Arg730*) [ 72 , 91 , 92 ]. Interestingly, sirtuins (SIRTs) are vital NAD + -dependent deacetylase enzymes that regulate autophagy [ 93 ], mitophagy [ 94 ], aging [ 95 ], and cytoskeletal (microtubule) functions [ 96 ] via dynamic changes in protein acetylation [ 54 ]. Similar to ADNP, SIRT1 regulates autophagy and mitochondrial functions [ 49 ], maintains genomic stability [ 45 ], enhances synaptic plasticity [ 46 ], suppresses inflammation [ 47 ], regulates cellular aging [ 48 ] and promotes neuroprotection [ 97 ]. Only recently, ADNP-SIRT1 interactions could be detected via WRD5 in a human neuroblastoma cell line [ 16 ], which could now be further validated in murine cerebellum modeling the died ADNP patient. Besides, we showed biochemical evidence for indirect binding of ADNP and SIRT1 via the microtubule end binding proteins (EB1/EB3), further linking autophagy, mitophagy, and cytoskeletal (microtubule) brain functions. Similarly, in other neurodevelopmental disorders such as Koolen-De Vries syndrome, dysfunctional autophagy was found to cause synaptic deficits in human IPSC cultures [ 79 , 98 ]. Our findings were further supported by gene set enrichment analysis of cellular stress responses [ 34 ] and mitochondrial activity-based assays which reveal disrupted mitochondrial gene expression via autophagy (mitophagy) processes and decreased mitochondrial activity in patient-derived fibroblasts and LCLs with an ADNP mutation. Interestingly, the ADNP-derived drug NAP (Davunetide) was shown to improve microtubule-dependent traffic, restore the autophagic flux and potentiate autophagosome-lysosome fusion, leading to autophagic vacuole clearance in Parkinson’s disease cells [ 99 ]. Finally, protein–protein interaction network analysis of our differential proteome analysis predicted SIRT1 in a central hub which links chromatin remodelers (e.g., ADNP, SMARCC2, HDAC2 and YY1) with autophagy signaling (BECN1, LAMP1 and LC3). Various chromatin remodeling proteins of the network have previously been ascribed as part of the ADNP-WDR5-SIRT1-BRG1-HDAC2-YY1 chromatin complex, which in part shares co-expression via transcriptional regulation [ 16 ].
In conclusion, our integrative multi-omics study for the first time has confirmed various ADNP mutant associated neurodevelopmental affected pathways at the epigenome-transcriptome-proteome level in primary brain tissue of an ADNP child, which previously have been described in in vitro LCL cultures and in vivo animal experiments substantiating strong cross-species and cross-cell type of molecular ADNP (disease) features. Moreover, our results hint towards a new functional link between the chromatin remodeler ADNP and the NAD + -deacetylase SIRT1 to control cytoskeletal and mitochondrial autophagy stress responses in neurodevelopment and plasticity. This novel molecular mechanism holds promise for new therapeutic strategies aimed at restoring mitochondrial (dys)function(s) in the Helsmoortel–Van der Aa syndrome.
Limitations of the study
In this study, we performed a molecular and biochemical autopsy study on the cerebellar tissue of a patient with Helsmoortel–Van der Aa syndrome, who has died after multiple organ failure following a second liver transplantation. We acknowledge several limitations that are either intrinsic to the study design or linked to the experimental course of action. First, we report the only post-mortem brain tissue that is currently available and compare it with an age-matched control subject. We acknowledge a gender difference between the ADNP patient (male) and control subject (female) but have consistently taken this difference into account during all bioinformatic analyses. In addition, we acknowledge that our control subject has a clinical diagnosis of Rett syndrome. However, we sequenced the entire ADNP gene, showing no genetic defect in the specimen. In addition, an expert pathologist examined brain sections using different immunohistochemical techniques, resulting no morphological abnormalities in the cerebellum. Although we are aware of the unique value of this material, we do acknowledge the lack of statistical power due to the limited sample size (n = 1). To overcome the lack of statistical power, we compared our findings of the human autopsy brain to several primary cell lines (e.g., LCLs and skin fibroblasts) derived from patients with different ADNP mutations and looked for gene expression similarities and dissimilarities across these tissues. Here, we observed a similar trend of gene expression changes in the human brain with in vitro and in vivo model systems, indicating a strong conservation of the ADNP gene and functioning. However, we report that this mutation is unique on its own, and no patient has been diagnosed with this ADNP mutation before. Therefore, comparisons to individuals with different ADNP mutations or genetically engineered mouse models cannot completely represent the molecular mechanisms underlying this specific mutation. On the topic of preservation of the autopsy material, we recognize a long post-mortem interval time i.e., the period of death of the patient and the time of the brain biopsy, which was 35-h after which liquid nitrogen was applied. To examine DNA and mRNA preservation in the cerebellum of our autopsy case, we subjected the extracted materials to the most sensitive bioanalyzer technologies to control for proper integrity. In addition, we also implemented very extensive validations e.g., pyrosequencing and RT-PCR for the implemented omics techniques using adequate controls. Similar to mRNA integrity, the preservation of proteins is not homogeneous since some molecules are more vulnerable than others [ 100 ]. In particular, we were unable to detect ADNP levels by means of western blotting in the cerebellum, but we did detect the protein in the cerebellum of the control subject. Since ADNP is already an unstable protein [ 7 , 29 ], the absence of the protein could potentially be attributed to degradation by active proteases in the brain during the post-mortem interval [ 101 ]. Lastly, we noted prominent changes in immune system-related pathways as indicated by our methylation, transcriptome, and proteome analyses of the autopsy brain. Although a subset of patients presented with thyroid problems [ 2 ] and recent investigations indicated a role of ADNP in T-helper cell differentiation [ 86 ], we show a critical attitude towards the interpretation of these results, as we cannot rule out certain bias in gene expression alterations following two liver transplantations and the administration of multiple antiepileptic drugs during the lifespan of this child with an ADNP mutation.
Availability of data and materials
The data presented in this study is available in the article or contained in the supplementary information. Correspondence and requests for material/raw data should be addressed to Claudio Peter D’Incal.
Vissers LELM, de Ligt J, Gilissen C et al (2010) A de novo paradigm for mental retardation. Nat Genet 42:1109–1112. https://doi.org/10.1038/ng.712
Article CAS PubMed Google Scholar
Helsmoortel C, Vulto-van Silfhout AT, Coe BP et al (2014) A SWI/SNF-related autism syndrome caused by de novo mutations in ADNP. Nat Genet 46:380–384. https://doi.org/10.1038/ng.2899
Article CAS PubMed PubMed Central Google Scholar
Vandeweyer G, Helsmoortel C, Van Dijck A et al (2014) The transcriptional regulator ADNP links the BAF (SWI/SNF) complexes with autism. Am J Med Genet C Semin Med Genet 166C:315–326. https://doi.org/10.1002/ajmg.c.31413
Van Dijck A, Vulto-van Silfhout AT, Cappuyns E et al (2019) Clinical presentation of a complex neurodevelopmental disorder caused by mutations in ADNP . Biol Psychiatry 85:287–297. https://doi.org/10.1016/j.biopsych.2018.02.1173
Article PubMed Google Scholar
Rolland T, Cliquet F, Anney RJL et al (2023) Phenotypic effects of genetic variants associated with autism. Nat Med 29:1671–1680. https://doi.org/10.1038/s41591-023-02408-2
Fu JM, Satterstrom FK, Peng M et al (2022) Rare coding variation provides insight into the genetic architecture and phenotypic context of autism. Nat Genet 54:1320–1331. https://doi.org/10.1038/s41588-022-01104-0
D’Incal CP, Van Rossem KE, De Man K et al (2023) Chromatin remodeler Activity-Dependent Neuroprotective Protein (ADNP) contributes to syndromic autism. Clin Epigenetics 15:45. https://doi.org/10.1186/s13148-023-01450-8
Pinhasov A, Mandel S, Torchinsky A et al (2003) Activity-dependent neuroprotective protein: a novel gene essential for brain formation. Brain Res Dev Brain Res 144:83–90. https://doi.org/10.1016/s0165-3806(03)00162-7
Mandel S, Gozes I (2007) Activity-dependent neuroprotective protein constitutes a novel element in the SWI/SNF chromatin remodeling complex. J Biol Chem 282:34448–34456. https://doi.org/10.1074/jbc.M704756200
Bassan M, Zamostiano R, Davidson A et al (1999) Complete sequence of a novel protein containing a femtomolar-activity-dependent neuroprotective peptide. J Neurochem 72:1283–1293. https://doi.org/10.1046/j.1471-4159.1999.0721283.x
Cappuyns E, Huyghebaert J, Vandeweyer G, Kooy RF (2018) Mutations in ADNP affect expression and subcellular localization of the protein. Cell Cycle 17:1068–1075. https://doi.org/10.1080/15384101.2018.1471313
Mosch K, Franz H, Soeroes S et al (2011) HP1 recruits activity-dependent neuroprotective protein to H3K9me3 marked pericentromeric heterochromatin for silencing of major satellite repeats. PLoS ONE 6:e15894. https://doi.org/10.1371/journal.pone.0015894
Kaaij LJT, Mohn F, van der Weide RH et al (2019) The ChAHP Complex Counteracts Chromatin Looping at CTCF Sites that Emerged from SINE Expansions in Mouse. Cell 178:1437-1451.e14. https://doi.org/10.1016/j.cell.2019.08.007
Sun X, Peng X, Cao Y et al (2020) ADNP promotes neural differentiation by modulating Wnt/β-catenin signaling. Nat Commun 11:2984. https://doi.org/10.1038/s41467-020-16799-0
Markenscoff-Papadimitriou E, Binyameen F, Whalen S et al (2021) Autism risk gene POGZ promotes chromatin accessibility and expression of clustered synaptic genes. Cell Rep 37:110089. https://doi.org/10.1016/j.celrep.2021.110089
Hadar A, Kapitansky O, Ganaiem M et al (2021) Introducing ADNP and SIRT1 as new partners regulating microtubules and histone methylation. Mol Psychiatry 26:6550–6561. https://doi.org/10.1038/s41380-021-01143-9
Mandel S, Rechavi G, Gozes I (2007) Activity-dependent neuroprotective protein (ADNP) differentially interacts with chromatin to regulate genes essential for embryogenesis. Dev Biol 303:814–824. https://doi.org/10.1016/j.ydbio.2006.11.039
Bend EG, Aref-Eshghi E, Everman DB et al (2019) Gene domain-specific DNA methylation episignatures highlight distinct molecular entities of ADNP syndrome. Clin Epigenetics 11:64. https://doi.org/10.1186/s13148-019-0658-5
Breen MS, Garg P, Tang L et al (2020) Episignatures stratifying ADNP syndrome show modest correlation with phenotype. BioRxiv. https://doi.org/10.1101/2020.04.01.014902
Article Google Scholar
Dingemans AJM, Hinne M, Truijen KMG et al (2023) PhenoScore quantifies phenotypic variation for rare genetic diseases by combining facial analysis with other clinical features using a machine-learning framework. Nat Genet 55:1598–1607. https://doi.org/10.1038/s41588-023-01469-w
Merenlender-Wagner A, Shemer Z, Touloumi O et al (2014) New horizons in schizophrenia treatment: autophagy protection is coupled with behavioral improvements in a mouse model of schizophrenia. Autophagy 10:2324–2332. https://doi.org/10.4161/15548627.2014.984274
Ivashko-Pachima Y, Sayas CL, Malishkevich A, Gozes I (2017) ADNP/NAP dramatically increase microtubule end-binding protein-Tau interaction: a novel avenue for protection against tauopathy. Mol Psychiatry 22:1335–1344. https://doi.org/10.1038/mp.2016.255
Oz S, Kapitansky O, Ivashco-Pachima Y et al (2014) The NAP motif of activity-dependent neuroprotective protein (ADNP) regulates dendritic spines through microtubule end binding proteins. Mol Psychiatry 19:1115–1124. https://doi.org/10.1038/mp.2014.97
Amram N, Hacohen-Kleiman G, Sragovich S et al (2016) Sexual divergence in microtubule function: the novel intranasal microtubule targeting SKIP normalizes axonal transport and enhances memory. Mol Psychiatry 21:1467–1476. https://doi.org/10.1038/mp.2015.208
Hacohen-Kleiman G, Sragovich S, Karmon G et al (2018) Activity-dependent neuroprotective protein deficiency models synaptic and developmental phenotypes of autism-like syndrome. J Clin Invest 128:4956–4969. https://doi.org/10.1172/JCI98199
Article PubMed PubMed Central Google Scholar
Karmon G, Sragovich S, Hacohen-Kleiman G et al (2022) Novel ADNP syndrome mice reveal dramatic sex-specific peripheral gene expression with brain synaptic and Tau pathologies. Biol Psychiatry 92:81–95. https://doi.org/10.1016/j.biopsych.2021.09.018
Ivashko-Pachima Y, Ganaiem M, Ben-Horin-Hazak I et al (2022) SH3- and actin-binding domains connect ADNP and SHANK3, revealing a fundamental shared mechanism underlying autism. Mol Psychiatry 27:3316–3327. https://doi.org/10.1038/s41380-022-01603-w
Dresner E, Malishkevich A, Arviv C et al (2012) Novel evolutionary-conserved role for the activity-dependent neuroprotective protein (ADNP) family that is important for erythropoiesis. J Biol Chem 287:40173–40185. https://doi.org/10.1074/jbc.M112.387027
D’Incal CP, Cappuyns E, Choukri K et al (2022) In search of the hidden protein: optimization of detection strategies for autism-associated activity-dependent neuroprotective protein (ADNP) mutants. Res Sq. https://doi.org/10.21203/rs.3.rs-1954095/v1
Aryee MJ, Jaffe AE, Corrada-Bravo H et al (2014) Minfi: a flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 30:1363–1369. https://doi.org/10.1093/bioinformatics/btu049
Zhou Y, Zhou B, Pache L et al (2019) Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun 10:1523. https://doi.org/10.1038/s41467-019-09234-6
Akhmedov M, Martinelli A, Geiger R, Kwee I (2020) Omics playground: a comprehensive self-service platform for visualization, analytics and exploration of Big Omics Data. NAR Genom Bioinform 2:lqz019. https://doi.org/10.1093/nargab/lqz019
Schindelin J, Arganda-Carreras I, Frise E et al (2012) Fiji: an open-source platform for biological-image analysis. Nat Methods 9:676–682. https://doi.org/10.1038/nmeth.2019
Bordi M, De Cegli R, Testa B et al (2021) A gene toolbox for monitoring autophagy transcription. Cell Death Dis 12:1044. https://doi.org/10.1038/s41419-021-04121-9
Zamostiano R, Pinhasov A, Gelber E et al (2001) Cloning and characterization of the human activity-dependent neuroprotective protein. J Biol Chem 276:708–714. https://doi.org/10.1074/jbc.M007416200
Mier P, Paladin L, Tamana S et al (2020) Disentangling the complexity of low complexity proteins. Brief Bioinform 21:458–472. https://doi.org/10.1093/bib/bbz007
Kastano K, Mier P, Andrade-Navarro MA (2021) The role of low complexity regions in protein interaction modes: an illustration in huntingtin. Int J Mol Sci. https://doi.org/10.3390/ijms22041727
Llamas B, Holland ML, Chen K et al (2012) High-resolution analysis of cytosine methylation in ancient DNA. PLoS ONE 7:e30226. https://doi.org/10.1371/journal.pone.0030226
Ostapcuk V, Mohn F, Carl SH et al (2018) Activity-dependent neuroprotective protein recruits HP1 and CHD4 to control lineage-specifying genes. Nature 557:739–743. https://doi.org/10.1038/s41586-018-0153-8
Tarazona S, Furió-Tarí P, Turrà D et al (2015) Data quality aware analysis of differential expression in RNA-seq with NOISeq R/Bioc package. Nucleic Acids Res 43:e140. https://doi.org/10.1093/nar/gkv711
Carlyle BC, Kitchen RR, Kanyo JE et al (2017) A multiregional proteomic survey of the postnatal human brain. Nat Neurosci 20:1787–1795. https://doi.org/10.1038/s41593-017-0011-2
Wang Y-M, Qiu M-Y, Liu Q et al (2021) Critical role of dysfunctional mitochondria and defective mitophagy in autism spectrum disorders. Brain Res Bull 168:138–145. https://doi.org/10.1016/j.brainresbull.2020.12.022
Hollis F, Kanellopoulos AK, Bagni C (2017) Mitochondrial dysfunction in Autism Spectrum Disorder: clinical features and perspectives. Curr Opin Neurobiol 45:178–187. https://doi.org/10.1016/j.conb.2017.05.018
Balachandar V, Rajagopalan K, Jayaramayya K et al (2021) Mitochondrial dysfunction: A hidden trigger of autism? Genes Dis 8:629–639. https://doi.org/10.1016/j.gendis.2020.07.002
Gonfloni S, Iannizzotto V, Maiani E et al (2014) P53 and Sirt1: routes of metabolism and genome stability. Biochem Pharmacol 92:149–156. https://doi.org/10.1016/j.bcp.2014.08.034
Michán S, Li Y, Chou MM-H et al (2010) SIRT1 is essential for normal cognitive function and synaptic plasticity. J Neurosci 30:9695–9707. https://doi.org/10.1523/JNEUROSCI.0027-10.2010
Xie J, Zhang X, Zhang L (2013) Negative regulation of inflammation by SIRT1. Pharmacol Res 67:60–67. https://doi.org/10.1016/j.phrs.2012.10.010
Godoy JA, Zolezzi JM, Braidy N, Inestrosa NC (2014) Role of Sirt1 during the ageing process: relevance to protection of synapses in the brain. Mol Neurobiol 50:744–756. https://doi.org/10.1007/s12035-014-8645-5
Ou X, Lee MR, Huang X et al (2014) SIRT1 positively regulates autophagy and mitochondria function in embryonic stem cells under oxidative stress. Stem Cells 32:1183–1194. https://doi.org/10.1002/stem.1641
Wang G, Fu Y, Hu F et al (2017) Loss of BRG1 induces CRC cell senescence by regulating p53/p21 pathway. Cell Death Dis 8:e2607. https://doi.org/10.1038/cddis.2017.1
Jiang K, Toedt G, Montenegro Gouveia S et al (2012) A Proteome-wide screen for mammalian SxIP motif-containing microtubule plus-end tracking proteins. Curr Biol 22:1800–1807. https://doi.org/10.1016/j.cub.2012.07.047
Almeida TB, Carnell AJ, Barsukov IL, Berry NG (2017) Targeting SxIP-EB1 interaction: an integrated approach to the discovery of small molecule modulators of dynamic binding sites. Sci Rep 7:15533. https://doi.org/10.1038/s41598-017-15502-6
Merenlender-Wagner A, Malishkevich A, Shemer Z et al (2015) Autophagy has a key role in the pathophysiology of schizophrenia. Mol Psychiatry 20:126–132. https://doi.org/10.1038/mp.2013.174
Salminen A, Kaarniranta K (2009) SIRT1: regulation of longevity via autophagy. Cell Signal 21:1356–1360. https://doi.org/10.1016/j.cellsig.2009.02.014
Vulih-Shultzman I, Pinhasov A, Mandel S et al (2007) Activity-dependent neuroprotective protein snippet NAP reduces tau hyperphosphorylation and enhances learning in a novel transgenic mouse model. J Pharmacol Exp Ther 323:438–449. https://doi.org/10.1124/jpet.107.129551
Xu J, Jackson CW, Khoury N et al (2018) Brain SIRT1 mediates metabolic homeostasis and neuroprotection. Front Endocrinol (Lausanne) 9:702. https://doi.org/10.3389/fendo.2018.00702
Chuang Y-C, Chen S-D, Jou S-B et al (2019) Sirtuin 1 regulates mitochondrial biogenesis and provides an endogenous neuroprotective mechanism against seizure-induced neuronal cell death in the hippocampus following status epilepticus. Int J Mol Sci. https://doi.org/10.3390/ijms20143588
Zhang X, Chen S, Huang K, Le W (2013) Why should autophagic flux be assessed? Acta Pharmacol Sin 34:595–599. https://doi.org/10.1038/aps.2012.184
Mizushima N, Murphy LO (2020) Autophagy assays for biological discovery and therapeutic development. Trends Biochem Sci 45:1080–1093. https://doi.org/10.1016/j.tibs.2020.07.006
Gozes I, Van Dijck A, Hacohen-Kleiman G et al (2017) Premature primary tooth eruption in cognitive/motor-delayed ADNP-mutated children. Transl Psychiatry 7:e1043. https://doi.org/10.1038/tp.2017.27
Bennison SA, Blazejewski SM, Liu X et al (2023) The cytoplasmic localization of ADNP through 14–3-3 promotes sex-dependent neuronal morphogenesis, cortical connectivity, and calcium signaling. Mol Psychiatry 28:1946–1959. https://doi.org/10.1038/s41380-022-01939-3
Arjun McKinney A, Petrova R, Panagiotakos G (2022) Calcium and activity-dependent signaling in the developing cerebral cortex. Development. https://doi.org/10.1242/dev.198853
Miller DE, Squire A, Bennett JT (2020) A child with autism, behavioral issues, and dysmorphic features found to have a tandem duplication within CTNND2 by mate-pair sequencing. Am J Med Genet A 182:543–547. https://doi.org/10.1002/ajmg.a.61442
Belcaro C, Dipresa S, Morini G et al (2015) CTNND2 deletion and intellectual disability. Gene 565:146–149. https://doi.org/10.1016/j.gene.2015.03.054
Ligon C, Seong E, Schroeder EJ et al (2020) δ-Catenin engages the autophagy pathway to sculpt the developing dendritic arbor. J Biol Chem 295:10988–11001. https://doi.org/10.1074/jbc.RA120.013058
Duan D, Fu Y, Paxinos G, Watson C (2013) Spatiotemporal expression patterns of Pax6 in the brain of embryonic, newborn, and adult mice. Brain Struct Funct 218:353–372. https://doi.org/10.1007/s00429-012-0397-2
Xu R, Brawner AT, Li S et al (2019) OLIG2 drives abnormal neurodevelopmental phenotypes in human iPSC-based organoid and chimeric mouse models of down syndrome. Cell Stem Cell 24:908-926.e8. https://doi.org/10.1016/j.stem.2019.04.014
Kanwore K, Guo X-X, Abdulrahman AA et al (2021) SOX1 is a backup gene for brain neurons and glioma stem cell protection and proliferation. Mol Neurobiol 58:2634–2642. https://doi.org/10.1007/s12035-020-02240-6
Bernal A, Arranz L (2018) Nestin-expressing progenitor cells: function, identity and therapeutic implications. Cell Mol Life Sci 75:2177–2195. https://doi.org/10.1007/s00018-018-2794-z
Boyl PP, Signore M, Annino A et al (2001) Otx genes in the development and evolution of the vertebrate brain. Int J Dev Neurosci 19:353–363. https://doi.org/10.1016/s0736-5748(01)00003-x
Lee HHC, Bernard C, Ye Z et al (2017) Genetic Otx2 mis-localization delays critical period plasticity across brain regions. Mol Psychiatry 22:680–688. https://doi.org/10.1038/mp.2017.1
Grigg I, Ivashko-Pachima Y, Hait TA et al (2020) Tauopathy in the young autistic brain: novel biomarker and therapeutic target. Transl Psychiatry 10:228. https://doi.org/10.1038/s41398-020-00904-4
Sragovich S, Malishkevich A, Piontkewitz Y et al (2019) The autism/neuroprotection-linked ADNP/NAP regulate the excitatory glutamatergic synapse. Transl Psychiatry 9:2. https://doi.org/10.1038/s41398-018-0357-6
Yu M, Liu Y, Wang Y et al (2020) Epithelial wnt10a is essential for tooth root furcation morphogenesis. J Dent Res 99:311–319. https://doi.org/10.1177/0022034519897607
Ming Y, Deng Z, Tian X et al (2022) Methyltransferase METTL3 reduces hippocampal neuron apoptosis in a mouse model of autism through the MALAT1/SFRP2/Wnt/β-catenin Axis. Psychiatry Investig 19:771–787. https://doi.org/10.30773/pi.2021.0370
Zhang F, Kang Y, Wang M et al (2018) Fragile X mental retardation protein modulates the stability of its m6A-marked messenger RNA targets. Hum Mol Genet 27:3936–3950. https://doi.org/10.1093/hmg/ddy292
Park S-Y, Avraham HK, Avraham S (2004) RAFTK/Pyk2 activation is mediated by trans-acting autophosphorylation in a Src-independent manner. J Biol Chem 279:33315–33322. https://doi.org/10.1074/jbc.M313527200
Sragovich S, Merenlender-Wagner A, Gozes I (2017) ADNP plays a key role in autophagy: from autism to schizophrenia and Alzheimer’s disease. BioEssays. https://doi.org/10.1002/bies.201700054
Lewerissa EI, Nadif Kasri N, Linda K (2024) Epigenetic regulation of autophagy-related genes: implications for neurodevelopmental disorders. Autophagy 20:15–28. https://doi.org/10.1080/15548627.2023.2250217
Wengrod J, Martin L, Wang D et al (2013) Inhibition of nonsense-mediated RNA decay activates autophagy. Mol Cell Biol 33:2128–2135. https://doi.org/10.1128/MCB.00174-13
Jolly LA, Homan CC, Jacob R et al (2013) The UPF3B gene, implicated in intellectual disability, autism, ADHD and childhood onset schizophrenia regulates neural progenitor cell behaviour and neuronal outgrowth. Hum Mol Genet 22:4673–4687. https://doi.org/10.1093/hmg/ddt315
Huang L, Shum EY, Jones SH et al (2018) A Upf3b-mutant mouse model with behavioral and neurogenesis defects. Mol Psychiatry 23:1773–1786. https://doi.org/10.1038/mp.2017.173
Domingo D, Nawaz U, Corbett M et al (2020) A synonymous UPF3B variant causing a speech disorder implicates NMD as a regulator of neurodevelopmental disorder gene networks. Hum Mol Genet 29:2568–2578. https://doi.org/10.1093/hmg/ddaa151
Nakamura S, Oba M, Suzuki M et al (2019) Suppression of autophagic activity by Rubicon is a signature of aging. Nat Commun 10:847. https://doi.org/10.1038/s41467-019-08729-6
Gozes I (2016) The cytoskeleton as a drug target for neuroprotection: the case of the autism-mutated ADNP. Biol Chem 397:177–184. https://doi.org/10.1515/hsz-2015-0152
Ferreira ACF, Szeto ACH, Clark PA et al (2023) Neuroprotective protein ADNP-dependent histone remodeling complex promotes T helper 2 immune cell differentiation. Immunity 56:1468-1484.e7. https://doi.org/10.1016/j.immuni.2023.05.010
Rose S, Niyazov DM, Rossignol DA et al (2018) Clinical and molecular characteristics of mitochondrial dysfunction in autism spectrum disorder. Mol Diagn Ther 22:571–593. https://doi.org/10.1007/s40291-018-0352-x
Rossignol DA, Frye RE (2012) Mitochondrial dysfunction in autism spectrum disorders: a systematic review and meta-analysis. Mol Psychiatry 17:290–314. https://doi.org/10.1038/mp.2010.136
Lim A, Thomas RH (2020) The mitochondrial epilepsies. Eur J Paediatr Neurol 24:47–52. https://doi.org/10.1016/j.ejpn.2019.12.021
Chang S-J, Yu B-C (2010) Mitochondrial matters of the brain: mitochondrial dysfunction and oxidative status in epilepsy. J Bioenerg Biomembr 42:457–459. https://doi.org/10.1007/s10863-010-9317-4
Mollinedo P, Kapitansky O, Gonzalez-Lamuño D et al (2019) Cellular and animal models of skin alterations in the autism-related ADNP syndrome. Sci Rep 9:736. https://doi.org/10.1038/s41598-018-36859-2
Gozes I, Ivashko-Pachima Y, Kapitansky O et al (2019) Single-cell analysis of cytoskeleton dynamics: from isoelectric focusing to live cell imaging and RNA-seq. J Neurosci Methods 323:119–124. https://doi.org/10.1016/j.jneumeth.2019.05.014
Li G-Z, Randev RK, Soeriyadi AH et al (2010) Investigation into thiol-(meth)acrylate Michael addition reactions using amine and phosphine catalysts. Polym Chem 1:1196. https://doi.org/10.1039/c0py00100g
Article CAS Google Scholar
Aventaggiato M, Vernucci E, Barreca F et al (2021) Sirtuins’ control of autophagy and mitophagy in cancer. Pharmacol Ther 221:107748. https://doi.org/10.1016/j.pharmthera.2020.107748
Wan W, Hua F, Fang P et al (2022) Regulation of mitophagy by sirtuin family proteins: a vital role in aging and age-related diseases. Front Aging Neurosci 14:845330. https://doi.org/10.3389/fnagi.2022.845330
Motonishi S, Nangaku M, Wada T et al (2015) Sirtuin1 maintains actin cytoskeleton by deacetylation of cortactin in injured podocytes. J Am Soc Nephrol 26:1939–1959. https://doi.org/10.1681/ASN.2014030289
Gomes BAQ, Silva JPB, Romeiro CFR et al (2018) Neuroprotective mechanisms of resveratrol in alzheimer’s disease: role of SIRT1. Oxid Med Cell Longev 2018:8152373. https://doi.org/10.1155/2018/8152373
Linda K, Lewerissa EI, Verboven AHA et al (2022) Imbalanced autophagy causes synaptic deficits in a human model for neurodevelopmental disorders. Autophagy 18:423–442. https://doi.org/10.1080/15548627.2021.1936777
Esteves AR, Gozes I, Cardoso SM (2014) The rescue of microtubule-dependent traffic recovers mitochondrial function in Parkinson’s disease. Biochim Biophys Acta 1842:7–21. https://doi.org/10.1016/j.bbadis.2013.10.003
Ferrer I, Martinez A, Boluda S et al (2008) Brain banks: benefits, limitations and cautions concerning the use of post-mortem brain tissue for molecular studies. Cell Tissue Bank 9:181–194. https://doi.org/10.1007/s10561-008-9077-0
Gozes I, Shazman S (2022) STOP codon mutations at sites of natural caspase cleavage are implicated in autism and Alzheimer’s disease: the case of ADNP. Front Endocrinol 13:364
Download references
Acknowledgements
We are profoundly grateful for the family to support our research study. We would especially like to thank our great interns Kirsty Szrama, Kaoutar Choukri, Amber Buys, and Kim Van Meel for their motivation and help during this project as partial fulfillment of their thesis. We thank Olivia Beck and Gabriela Munteanu for expertise and assistance with the lymphoblastoid cell lines. We express our appreciation towards Bart De Vil and Patrick Cras (IBB-Neurobiobank, Antwerp University) for providing us with post-mortem control cerebellar tissue. We are grateful for the help and expertise of Sofie Theys and Dr. Isabel Pintelon regarding the immunohistochemical stainings (ACAM, Antwerp University). We thank the Laboratory of Microbiology, Parasitology and Hygiene (LMPH) of Prof. dr. Guy Caljon for the training and use of the Tecan Spark machine. We thank the lovely and helpful Michelle Zurek (Laboratory of Physiopharmacology) for donating the Bafilomycin A1 compound to assess the autophagic flux in the ADNP cell lines, as well as for the help with interpretation of the results. Ultimately, we thank the ADNP Kids Research Foundation for continuous collaboration.
R. Frank Kooy acknowledges the support of the Research Fund of the University of Antwerp OEC-Methusalem grant “GENOMED”. This work was in part financed by grants from the ERA-NET NEURON “ADNPinMED”. This article is also based upon work from COST Action International Nucleome Consortium (INC) CA18127, supported by COST (European Cooperation in Science and Technology) ascribed to Wim Vanden Berghe.
Author information
Authors and affiliations.
Department of Medical Genetics, University of Antwerp, Prins Boudewijnlaan 43/6, 2650, Edegem, Antwerp, Belgium
Claudio D’Incal, Anke Van Dijck, Joe Ibrahim, Kevin De Man, Ellen Elinck, Nathalie Van der Aa, Ligia Mateiu & R. Frank Kooy
Protein Chemistry, Proteomics and Epigenetic Signaling (PPES), Epigenetic Signaling lab (PPES), Department of Biomedical Sciences, University of Antwerp, Universiteitsplein 1, 2610, Wilrijk, Antwerp, Belgium
Claudio D’Incal, Kevin De Man, Lina Bastini, Anthony Konings & Wim Vanden Berghe
Family Medicine and Population Health (FAMPOP), Department of Medicine and Health Sciences, University of Antwerp, Antwerp, Belgium
Anke Van Dijck
The Elton Laboratory for Molecular Neuroendocrinology, Department of Human Molecular Genetics and Biochemistry, Sackler Faculty of Medicine, Adams Super Center for Brain Studies and Sagol School of Neuroscience, Tel Aviv University, Tel Aviv, Israel
lllana Gozes
Clinical Department of Pathology and Cytology, University Hospital Centre Zagreb, Zagreb, Croatia
Zlatko Marusic
Division of Pediatric Gastroenterology, Hepatology and Nutrition, Department of Pediatrics, School of Medicine, University Hospital Centre Zagreb, Zagreb, Croatia
Mirna Anicic & Jurica Vukovic
You can also search for this author in PubMed Google Scholar
Contributions
C.P.D. performed the experiments, conceptualized the experimental design, performed data analysis, wrote the manuscript text, and prepared all the figures. A.V.D. is the neurologist who collected the clinical data. J.I. provided the bioinformatic analysis of the methylation data. K.D.M., L.B., and A.K. were active participants in the experiments under supervision of C.D., E.E. conducted all the gene expression assays using RT-PCR under supervision of C.D., I.G. was involved in the tissue collection of the autopsy and reviewed the manuscript. Z.M. provided the postmortem tissue. M.A. and J.V. also provided the clinical data together with A.V.D., N.V.D.A. is a pediatrician and responsible for obtaining the LCLs from her patients. L.M. provided the bioinformatic analysis of the RNA sequencing of the brain and LCLs. W.V.B. reviewed the manuscript. R.F.K. reviewed the manuscript. All authors reviewed the manuscript.
Corresponding authors
Correspondence to Wim Vanden Berghe or R. Frank Kooy .
Ethics declarations
Ethics approval and consent to participate.
Experiments including C57BL6/JCr mice were in compliance with the EU Directive 2010/63/EU under ECD code 2022–59 after approval by the Animal Ethics Committee of the University of Antwerp. The cerebellar tissue of a died nine-year-old child was a kind gift from the Institute Born-Bunge vzw IBB NeuroBioBank of the University of Antwerp under HMTA20210040 by the Ethics Committee of the Hospital Network Antwerp (ZNA) Middelheim and HogeBeuken, Antwerp, Belgium. Clinical information of a died six-year-old male carrying the heterozygous c.1676dupA/p.His559Glnfs*3 ADNP mutation was received under informed consent of the parents under B300201627322 and approved by the Ethics Committee of the Antwerp University Hospital, Antwerp, Belgium. HEK293T cells were purchased from ATCC at a low passage number with consistent use of cells below a passage number of 15. LCLs were generated from peripheral blood lymphocytes donated by consenting patients and guardians/physicians in the laboratory of Frank Kooy. Immortalized skin fibroblasts were generated by consenting patients and guardians/physicians in the laboratory of Frank Kooy.
Consent for publication
An written informed consent under for publication died six-year-old male carrying the heterozygous c.1676dupA/p.His559Glnfs*3 ADNP mutation was received by the mother (caregiver) and gives permission to the medical doctors of the Department of Medical Genetics, University Hospital Antwerp, to use photographs, biopsies and/or X-rays for medical reasons (discussion between colleagues, teaching, or publication in scientific papers). The Institute Born-Bunge vzw IBB NeuroBioBank of the University of Antwerp acknowledges publication in all oral presentations, written or press related releases related to the use of the cerebellar tissue of a died nine-year-old female. All patient-derived cell lines were obtained under a written informed consent for publication by the patients, their guardians, and the tending clinicians.
Accordance to ARRIVE guidelines
We confirm that this study is reported in accordance with the ARRIVE guidelines ( https://arriveguidelines.org ).
Competing interests
Professor Dr. Em. Illana Gozes serves as VP drug development at ExoNavis Therapeutics LTD, clinically developing Davunetide.
Additional information
Publisher's note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
: Table S1 . Specifications of the human subjects, lymphoblastoid and fibroblastic cell lines. The table represents the anonymized patient IDs as a fictive number together with the WES-validated mutation in the ADNP gene. RNA purity determined by the 260/280 ratio and RIN integrity score of the RNA samples were also reported. Table S2 . Tested antibody overview. The table indicates the used antibodies for this study together with the manufacture and catalog number, host species, predicted reactivity, peptide sequence and the optimized dilution for each western blot experiment. Table S3 . Pyrosequencing primers. Table contains gene name, forward primer (5’ 3’), reverse primer (5’ 3’), location of the biotin tag of either the forward (Fwd) or reverse (Rev) primer, sequencing primer (5’ 3’), nucleotide sequence for CpG analysis and predication of the EPIC Beadchip array result. Table S4 . RT-PCR primer sequences. The table represent the forward and reverse primer sequences (5’ 3’) for expression analysis of ADNP, brain and lymphoblastoid transcriptome, and mitochondrial gene panel confirmations. Table S5 . Mitophagy-related gene panel using for screening the RNA sequencing data of ADNP lymphoblastoid cell lines (LCLs). Table S6 . Phenotype and clinical features of the post-mortem ADNP patient carrying the c.1676duplA/p.His559Glnfs*3 mutation in comparison with a cohort study by Van Dijck et al. ( 2019 ) entailing 78 HVDAS individuals.
Additional file 2
: Human Infinium EPIC BeadChip Array data of the ADNP cerebellum as compared to an age-matched control subject . CpG probes with an absolute β-value > 0.1 (10% methylation differences) are represented.
Additional file 3
: Transcription factor motif analysis with Irregulon . Enriched transcription factor motifs in the ADNP cerebellum using the identified hypo- and hypermethylated genes. Motif enrichment was scored using the normalized enrichment score (NES).
Additional file 4
: List of differentially expressed genes in the ADNP cerebellum as compared to an age-matched control subject using the NOIseq non-parametric analysis package.
Additional file 5
: List of differentially expressed genes in the ADNP patient-derived lymphoblastoid cell lines as compared to age- and sex-matched control lymphoblastoid cell lines using the DESeq2 analysis package.
Additional file 6
: List of overlapping differentially expressed genes in the ADNP cerebellum that overlap with the differentially expressed genes in the ADNP patient-derived lymphoblastoid cell lines.
Additional file 7
: Correlation heatmap of the label-free quantification mass spectrometry (LFQ-MS) experiment in the ADNP cerebellum . Pairwise correlations of protein abundances characterizing the relationships between proteins of the global mass spectrometry experiment. Correlations between all proteins, clustering of protein groups and similarly behaving proteins are represented in the protein-protein correlation matrix calculated on the logarithmic intensities. A strong correlation between technical replicates (n = 5) was observed (red color), indicating high reproducibility as tested by the Pearson correlation. Negative correlations are represented in a blue color (see scaled bar legend).
Additional file 8
: List of differentially expressed proteins in the ADNP cerebellum (LFQ-MS).
Additional file 9
: Mitophagy transcriptomic gene signature in the ADNP cerebellum . Expression levels of mitophagy-related genes in the ADNP patient cerebellum compared to an age-matched control subject. mRNA sequencing demonstrated both an upregulated (red) as well as a downregulated (blue) mitophagy gene signature in the ADNP patient cerebellum as compared to an age-matched control subject.
Additional file 10
: Autophagic flux assessment in ADNP patient-derived lymphoblastoid cell lines using Bafilomycin A1 . A . The autophagic flux was determined in ADNP patient-derived and age- and sex-matched control lymphoblastoid cell lines by treatment with 160 nM of Bafilomycin A1 (BAF) for two hours. Protein extracts of untreated and BAF-treated cells were subjected to western blotting using anti-p62/SQSTM1 and anti-LC3 antibodies to assess the autophagic flux. Although p62 expression increased after BAF treatment (+BAF) compared to untreated (-BAF) cells, the difference was not significant in patients (PAT) and controls (CTR). However, the expression of LC3 significantly increased after BAF treatment and was significantly increased in PAT versus CTR post-treatment compared to untreated patient cells (PAT-BAF). All western blots were controlled by GAPDH to ensure equal loading. Image quantification was performed using ImageJ software. B . Graphical representation was performed in GraphPad Prism version 9.3.1 using a 2-way ANOVA with Sidak’s multiple comparisons test to assess the interaction of the genotype (PAT versus CTR) and treatment (-BAF versus +BAF).
Additional file 11
: Determination of the mitochondrial DNA copy number (mtDNA-CN) in ADNP patient-derived lymphoblastoid cells and fibroblastic cell lines.
Additional file 12
: Raw western blotting images.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ . The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Reprints and permissions
About this article
D’Incal, C., Van Dijck, A., Ibrahim, J. et al. ADNP dysregulates methylation and mitochondrial gene expression in the cerebellum of a Helsmoortel–Van der Aa syndrome autopsy case. acta neuropathol commun 12 , 62 (2024). https://doi.org/10.1186/s40478-024-01743-w
Download citation
Received : 23 November 2023
Accepted : 11 February 2024
Published : 18 April 2024
DOI : https://doi.org/10.1186/s40478-024-01743-w
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Activity-dependent neuroprotective protein (ADNP)
- Methylation
- Chromatin remodeler
- Mitochondria
- Post-mortem brain
- Helsmoortel–Van der Aa syndrome
- Sirtuin 1 (SIRT1)
- Find a journal
- Publish with us
- Track your research
IMAGES
VIDEO
COMMENTS
Introduction. Intellectual Disability is defined in DSM-5 as a conceptual, socially defective disorder that covers both intellectual and adaptive deficiencies and starts during the developmental stage. 1 Aggressive and challenging behaviors are frequently seen in these individuals with limited coping skills. 2 These behaviors, which are considered as the most important factors in applying to ...
Case Study of a Child with Intellectual Disability. Exclusively available on IvyPanda. Updated: Oct 29th, 2023. In this intellectual disability case study, the author looks at designing an education curriculum for Meagan, a 14-year-old student. We will write a custom essay on your topic.
accessible, accountable, and meaningful to the disability community. In this case study, adults with intellectual and developmental disabilities (IDD) contributed as co-researchers to a series of studies on mental health of adults with intellectual disability. The research model, specific engagement strategies, and lessons learned are shared.
Vicente and collegues (2019) established three separate panels comprising ten 'professionals' (paid-experts) six 'relatives' (unpaid-experts), and five people with intellectual disability. In this case the participation of people with intellectual disability was segregated, rather than modifying the study to facilitate full and equal ...
Intellectual disabilities affect 1-3% of the U.S. population. The Executive Vice President and Chief Clinical Officer for Prisma Health discusses how health care redesign should factor in this population to reduce the health disparities they face. Authors: Jonathan Gleason, MD, and Namita Seth Mohta, MD Author Info & Affiliations.
A double-baseline case study design explored the use of everyday technology applications and devices to support functional performance of three men aged 32, 33, and 55 years, with mild to moderate intellectual disabilities. Performance of selected tasks was video-recorded and analysed on four occasions for each participant.
Pert C, Jahoda A, Stenfert Kroese B, et al. (2013) Cognitive behavioural therapy from the perspective of clients with mild intellectual disabilities: a qualitative investigation of process issues. Journal of Intellectual Disability Research 57(4): 359-369. DOI: 10.1111/j.1365-2788.2012.01546.x
Of the 41 studies, 15 were quantitative studies, 12 were qualitative studies, six were mixed methods studies, and eight were case studies. 17 of the studies pertained to adults with a mild-to-moderate intellectual disability, seven studies pertained to adults with a moderate to severe intellectual disability, five studies pertained to adults ...
2.1 Instructional models for students with intellectual disabilities. The inclusion of students with ID has influenced the educational system to the extent that is has developed many instructional activities and teaching approaches specific to this type of student [16, 19, 34, 55].In some cases, the education of students with ID and students without ID is combined under an inclusive approach ...
The study established the special needs or weak areas and strengths of Chiunda a 9 year old girl with mild intellectual disability (IQ 55). Secondly, this study brought to light the limitations ...
A dramatherapy case study with a young man who has dual diagnosis of intellectual disability and mental health problems. The Arts in Psychotherapy, Vol. 50, Issue. , p. The Arts in Psychotherapy, Vol. 50, Issue. , p.
week from 8:15 am to 2:30 pm. The program offers skill building, physical and occupational. therapy, and pre-vocational training for individuals diagnosed with intellectual disabilities and. mental illness and who require moderate to high level of assistance. The environment.
6 hours for pain, Levothyroxin 50mcg PO daily. The task in this case is to assess the EENT status of a 45 year old individual with an intellectual disability using history-taking with patient. 18. Setting of Encounter: Primary Care Office SP: Seated on the exam room fully clothed.
Students with intellectual needs have problems with their cognitive capability and usually lack skills in social behavior and self-care (Jones & Goble, 2012). Even though there is a wide assortment in the cognitive ability of individuals with intellectual disabilities, they generally have an IQ of 70-75 or even lower (Gordon, 2018).
Background Studies have suggested that people with intellectual disability are disproportionately involved in crime both as perpetrators and victims. Method A case linkage design used three Australian contact-level databases, from disability services, public mental health services and police records. Rates of contact, and official records of victimisation and criminal charges were compared to ...
In the United States, students with mild intellectual disability ... In this qualitative case study, the researchers describe the teaching methods of a seventh grade, special education teacher who heavily emphasized visuals, such as diagrams and gestures, in her instructional approach. In her classroom, two students with MID demonstrated the ...
Ethical conflict may occur when laws and policies permit the physical restraint of a client with an intellectual disability even though this restraint use is contradictory to the official position held by social work organizations and the personally held beliefs of the social worker. For example, holding grips may be used by care staff to ...
A total of five subjects with moderate intellectual disability aged 7-12 years was selected for this study. The data were collected from eleven parents and teachers by interview. All data were ...
The purpose of the present study through the case study of a student with intellectual disabilities is to implement and evaluate the pedagogical tool, TISIPfSENs, in the secondary school microsystem.
Guardianship and Aging in Intellectual Disabilities Case Studies Part II. A companion piece to Guardianship I, which gives an overview of guardianship, this webinar demonstrates real world case examples. It reviews different situations and describes what is appropriate during certain circumstances and scenarios. The intended audience for this ...
In the present study, we describe a new patient affected by mild intellectual disability, speech disorder, and non-specific abnormalities on EEG and neuroimaging. Family studies identified a new de novo frameshift variant c.1818delG (p.(Gln606Hisfs*101)) in SATB1 .
The literature regarding social inclusion for people with an intellectual disability has been marked by inconsistency and lack of conceptual clarity (Amado et al., 2013; Overmars-Marx et al., 2014).For conceptual clarity, here we adopt the concept of social inclusion offered by Simplican et al. (2015), which proposed social inclusion as the interaction of two key life domains - interpersonal ...
Helsmoortel-Van der Aa syndrome is a neurodevelopmental disorder in which patients present with autism, intellectual disability, and frequent extra-neurological features such as feeding and gastrointestinal problems, visual impairments, and cardiac abnormalities. All patients exhibit heterozygous de novo nonsense or frameshift stop mutations in the Activity-Dependent Neuroprotective Protein ...